

The embryonic epicardium: an essential element of cardiac development
- PMID: 20477903
- PMCID: PMC3822997
- DOI: 10.1111/j.1582-4934.2010.01088.x
The embryonic epicardium: an essential element of cardiac development
Abstract
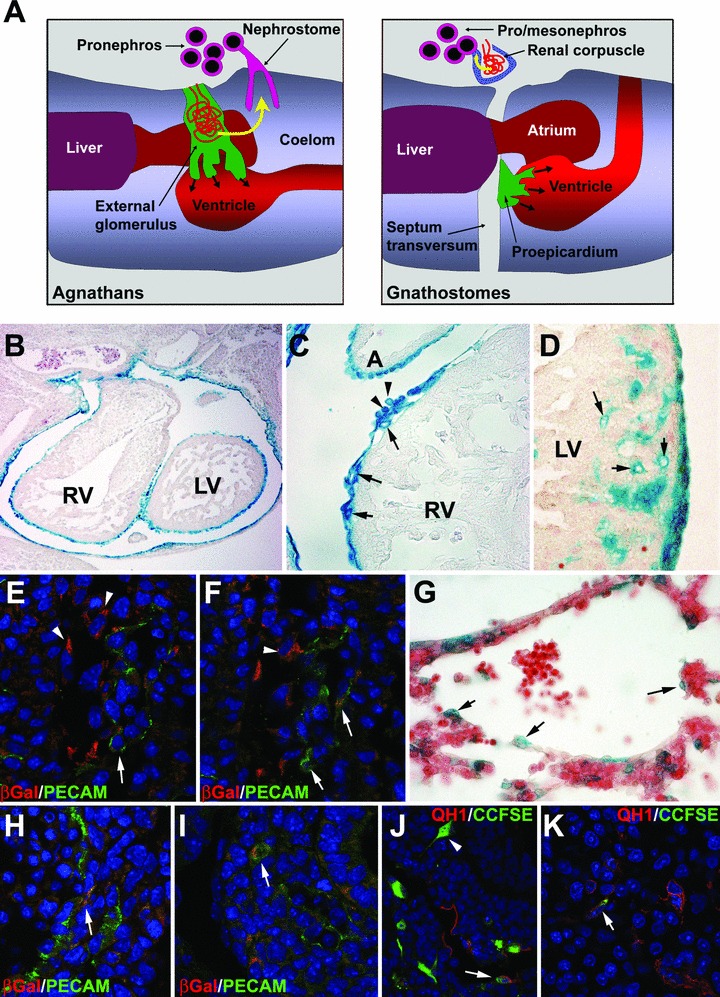
The epicardium has recently been identified as an active and essential element of cardiac development. Recent reports have unveiled a variety of functions performed by the embryonic epicardium, as well as the cellular and molecular mechanisms regulating them. However, despite its developmental importance, a number of unsolved issues related to embryonic epicardial biology persist. In this review, we will summarize our current knowledge about (i) the ontogeny and evolution of the epicardium, including a discussion on the evolutionary origins of the proepicardium (the epicardial primordium), (ii) the nature of epicardial-myocardial interactions during development, known to be essential for myocardial growth and maturation, and (iii) the contribution of epicardially derived cells to the vascular and connective tissue of the heart. We will finish with a note on the relationships existing between the primordia of the viscera and their coelomic epithelial lining. We would like to suggest that at least a part of the properties of the embryonic epicardium are shared by many other coelomic cell types, such that the role of epicardium in cardiac development is a particular example of a more general mechanism for the contribution of coelomic and coelomic-derived cells to the morphogenesis of organs such as the liver, kidneys, gonads or spleen.
© 2010 The Authors Journal compilation © 2010 Foundation for Cellular and Molecular Medicine/Blackwell Publishing Ltd.
Figures
References
-
- Pérez-Pomares JM, Macías D, García-Garrido L, et al. The origin of the subepicardial mesenchyme in the avian embryo, an immunohistochemical and quail-chick chimera study. Dev Biol. 1998;200:57–68. - PubMed
-
- Männer J, Pérez-Pomares JM, Macías D, et al. The origin, formation and developmental significance of the epicardium, a review. Cells Tiss Org. 2001;169:89–103. - PubMed
-
- Schulte I, Schlueter J, Abu-Issa R, et al. Morphological and molecular left-right asymmetries in the development of the proepicardium: a comparative analysis on mouse and chick embryos. Dev Dyn. 2007;236:684–95. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
