

The vomeronasal organ mediates interspecies defensive behaviors through detection of protein pheromone homologs
- PMID: 20478258
- PMCID: PMC2873972
- DOI: 10.1016/j.cell.2010.03.037
The vomeronasal organ mediates interspecies defensive behaviors through detection of protein pheromone homologs
Abstract
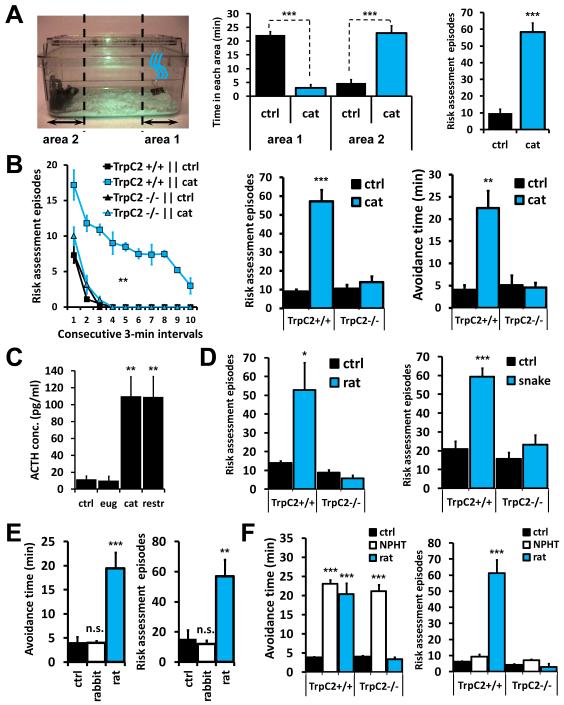
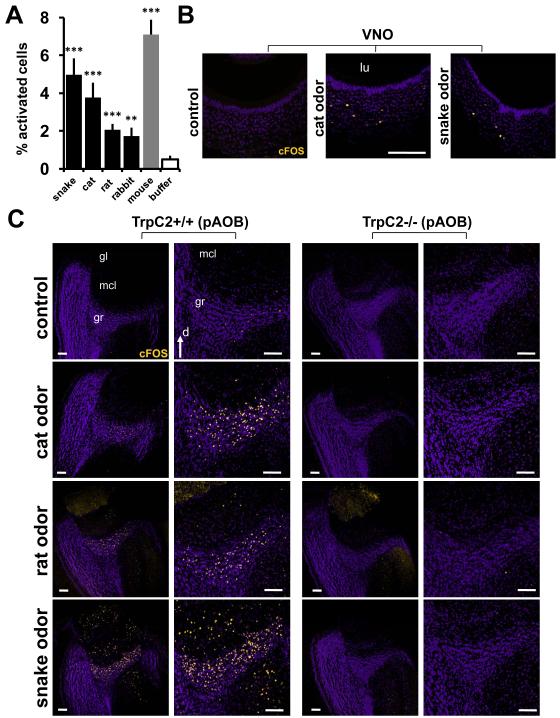
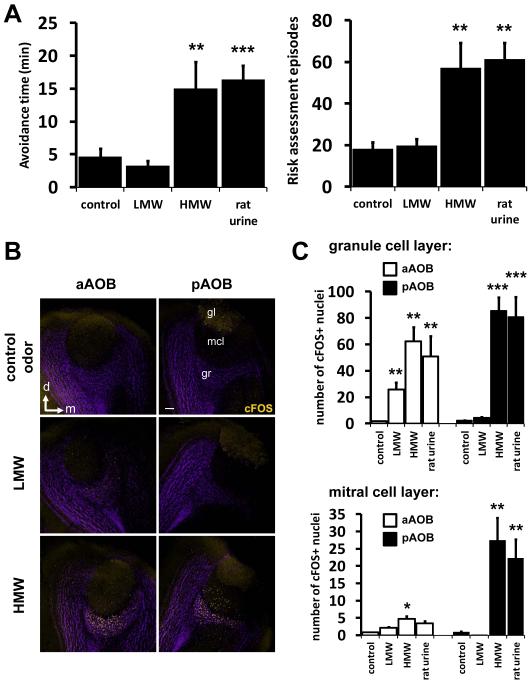
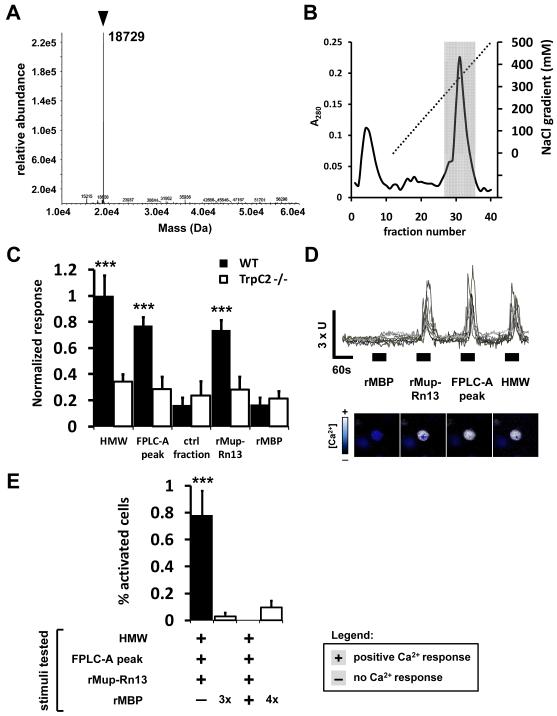
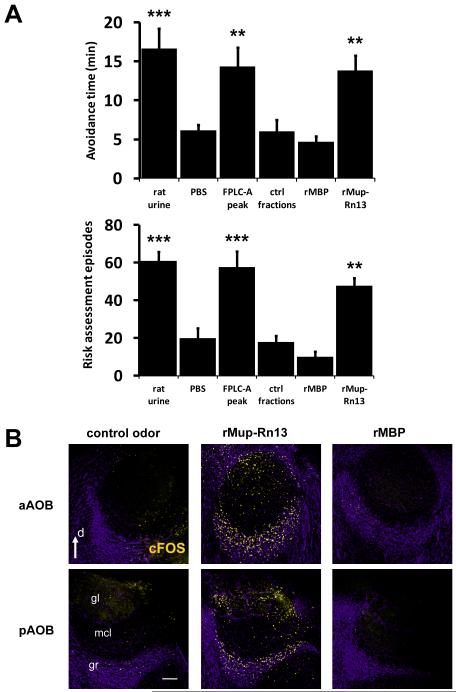
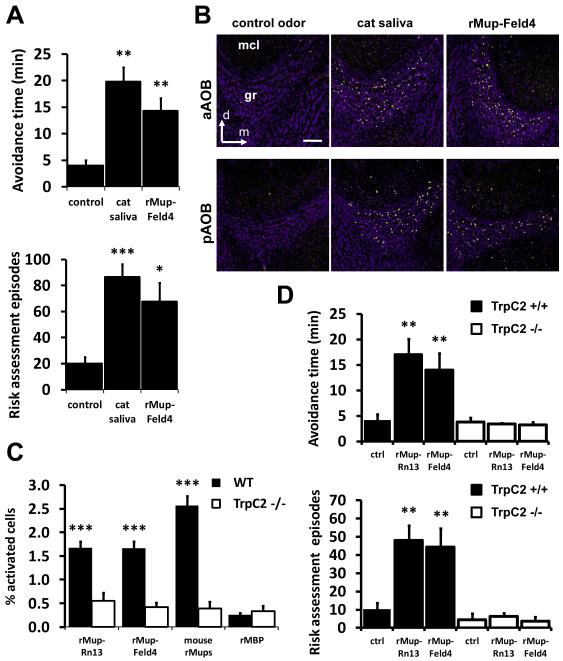
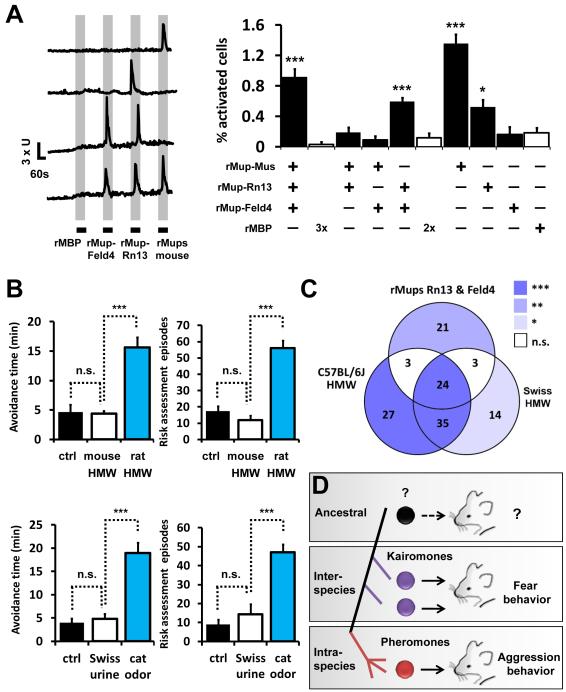
Potential predators emit uncharacterized chemosignals that warn receiving species of danger. Neurons that sense these stimuli remain unknown. Here we show that detection and processing of fear-evoking odors emitted from cat, rat, and snake require the function of sensory neurons in the vomeronasal organ. To investigate the molecular nature of the sensory cues emitted by predators, we isolated the salient ligands from two species using a combination of innate behavioral assays in naive receiving animals, calcium imaging, and c-Fos induction. Surprisingly, the defensive behavior-promoting activity released by other animals is encoded by species-specific ligands belonging to the major urinary protein (Mup) family, homologs of aggression-promoting mouse pheromones. We show that recombinant Mup proteins are sufficient to activate sensory neurons and initiate defensive behavior similarly to native odors. This co-option of existing sensory mechanisms provides a molecular solution to the difficult problem of evolving a variety of species-specific molecular detectors.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
The chemical MUPpeteer.Cell. 2010 May 14;141(4):568-70. doi: 10.1016/j.cell.2010.04.032. Cell. 2010. PMID: 20478249
References
-
- Apfelbach R, Blanchard CD, Blanchard RJ, Hayes RA, McGregor IS. The effects of predator odors in mammalian prey species: a review of field and laboratory studies. Neurosci Biobehav Rev. 2005;29:1123–1144. - PubMed
-
- Blanchard DC, Griebel G, Blanchard RJ. Mouse defensive behaviors: pharmacological and behavioral assays for anxiety and panic. Neurosci Biobehav Rev. 2001;25:205–218. - PubMed
-
- Buron G, Hacquemand R, Pourie G, Lucarz A, Jacquot L, Brand G. Comparative behavioral effects between synthetic 2,4,5-trimethylthiazoline (TMT) and the odor of natural fox (Vulpes vulpes) feces in mice. Behav Neurosci. 2007;121:1063–1072. - PubMed
-
- Chamero P, Marton TF, Logan DW, Flanagan K, Cruz JR, Saghatelian A, Cravatt BF, Stowers L. Identification of protein pheromones that promote aggressive behaviour. Nature. 2007;450:899–902. - PubMed
-
- Cheetham SA, Smith AL, Armstrong SD, Beynon RJ, Hurst JL. Limited variation in the major urinary proteins of laboratory mice. Physiol Behav. 2009;96:253–261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
