

Antiviral instruction of bone marrow leukocytes during respiratory viral infections
- PMID: 20478536
- PMCID: PMC2874206
- DOI: 10.1016/j.chom.2010.04.006
Antiviral instruction of bone marrow leukocytes during respiratory viral infections
Abstract
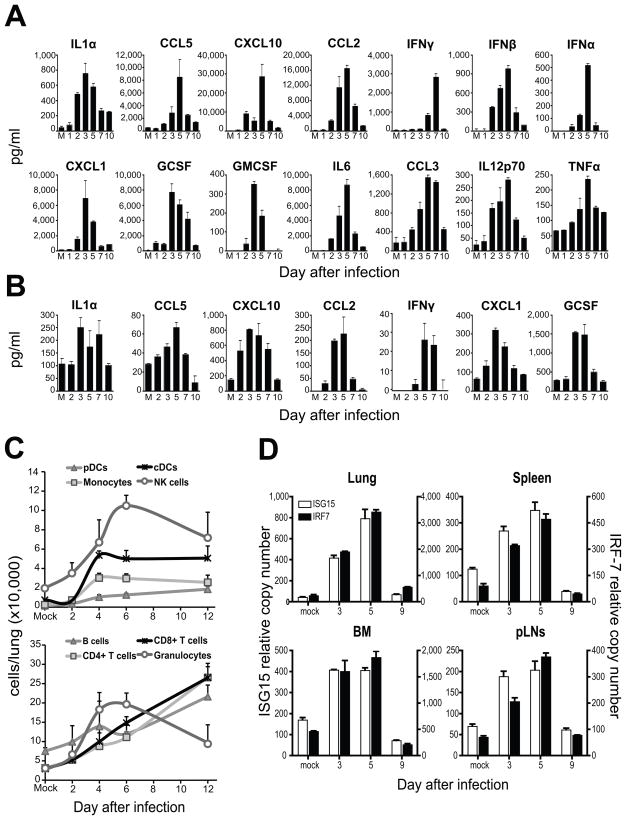
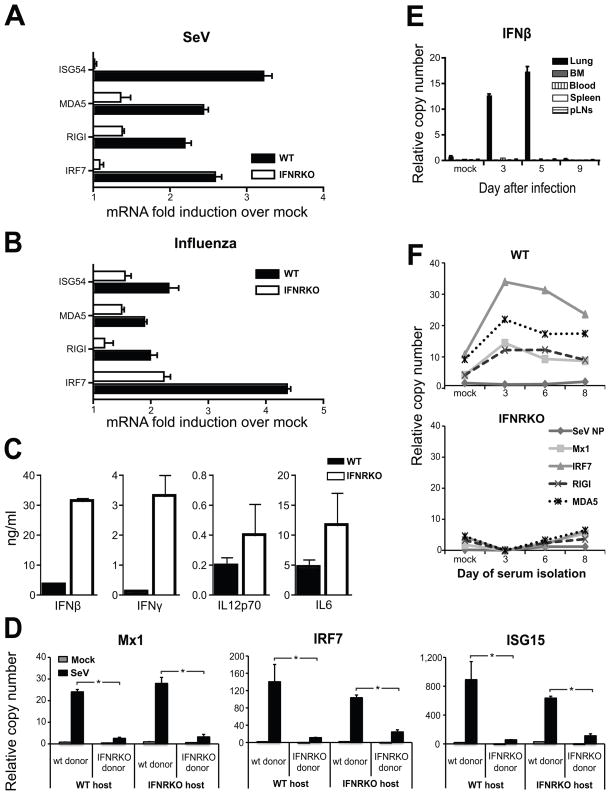
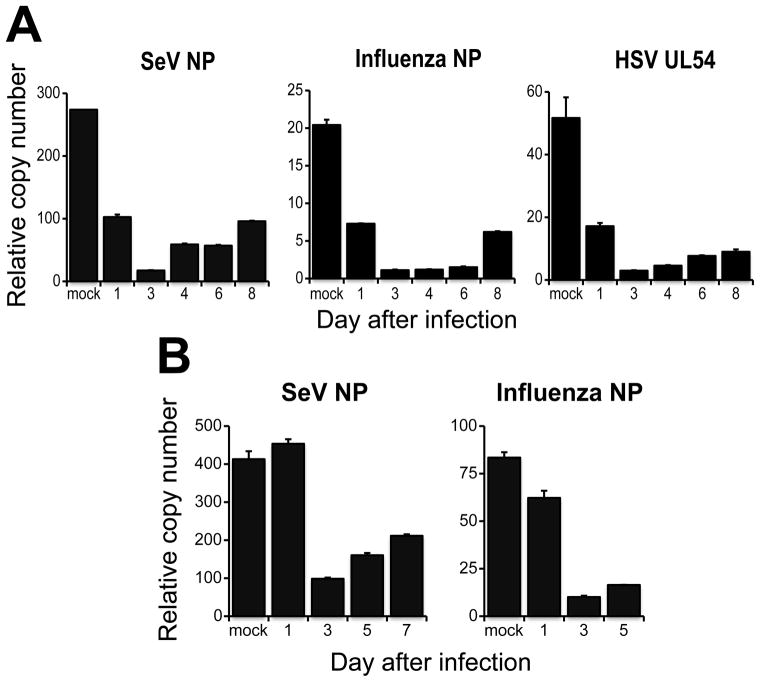
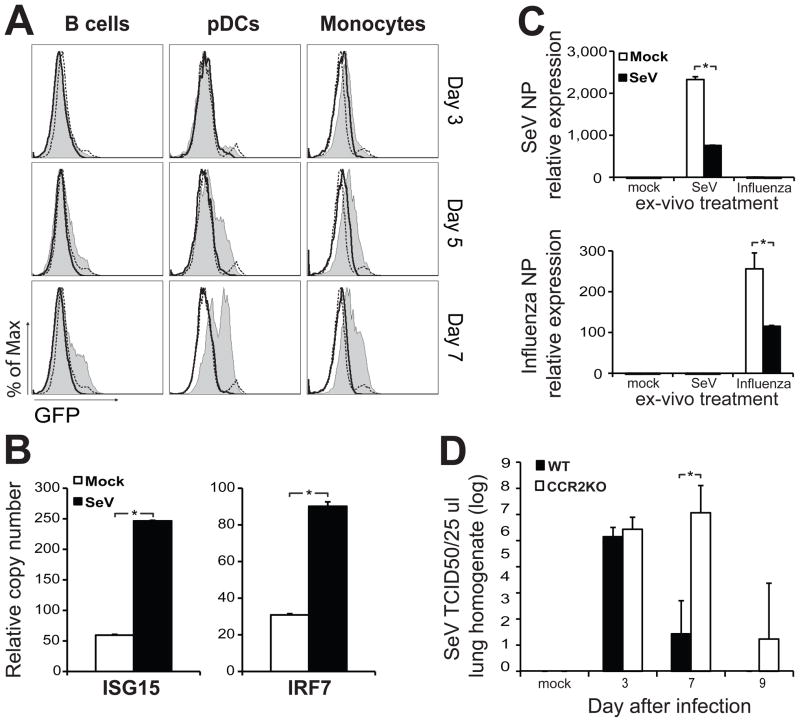
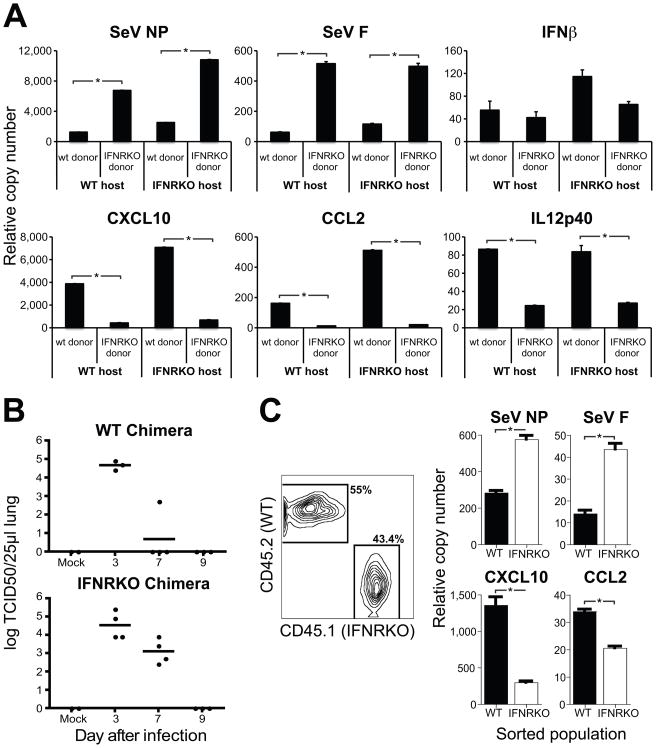
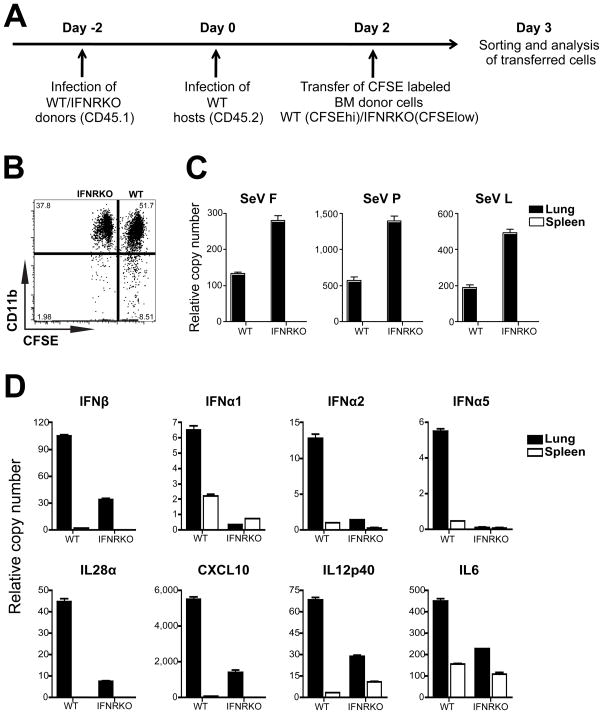
Respiratory viral infections trigger a robust inflammatory response in the lung, producing cytokines, chemokines, and growth factors that promote infiltration of effector leukocytes. Whereas the role of chemokines and infiltrating leukocytes in antiviral immunity is well studied, the effect that lung cytokines have on leukocytes in distal hematopoietic and lymphoid tissues and their role in antiviral immunity is unknown. We show that, during infection with influenza or Sendai virus, the lung communicates with the sterile bone marrow, the primary site of hematopoiesis, through type I interferons. While in the bone marrow, leukocytes exposed to type I interferons activate an antiviral transcriptional program and become resistant to infection with different viruses. The protected bone marrow leukocytes are capable of migrating to the infected lung and contribute to virus clearance. These findings show that appropriate instruction of cells during their development in the bone marrow is needed for effective control of infection.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A call to arms: interferons prepare bone marrow cells to battle peripheral infections.Cell Host Microbe. 2010 May 20;7(5):336-7. doi: 10.1016/j.chom.2010.05.004. Cell Host Microbe. 2010. PMID: 20478533
References
-
- Bernardini G, Sciume G, Bosisio D, Morrone S, Sozzani S, Santoni A. CCL3 and CXCL12 regulate trafficking of mouse bone marrow NK cell subsets. Blood. 2008;111:3626–3634. - PubMed
-
- Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Annu Rev Immunol. 1999;17:189–220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
