

Strigolactone regulation of shoot branching in chrysanthemum (Dendranthema grandiflorum)
- PMID: 20478970
- PMCID: PMC2892150
- DOI: 10.1093/jxb/erq133
Strigolactone regulation of shoot branching in chrysanthemum (Dendranthema grandiflorum)
Abstract
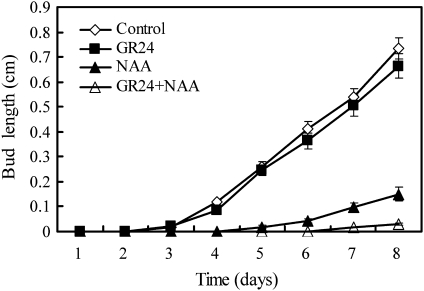
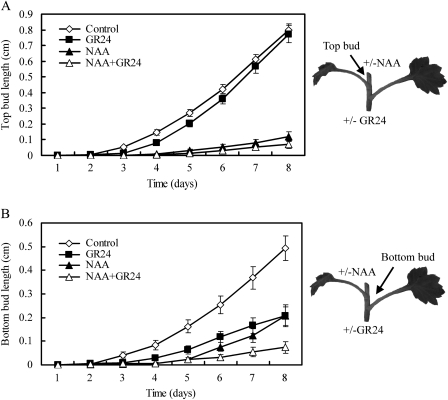
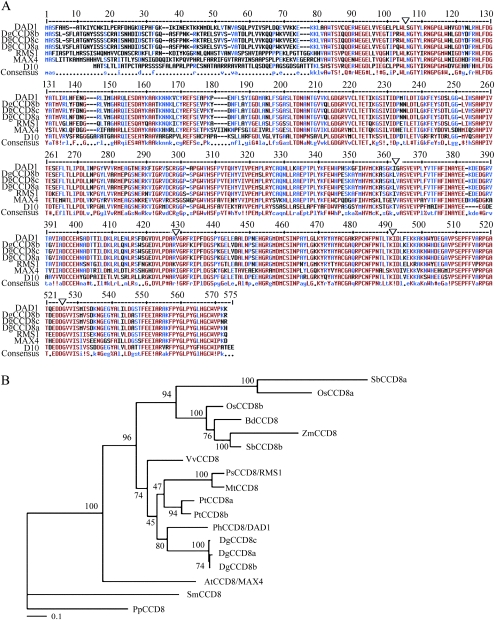
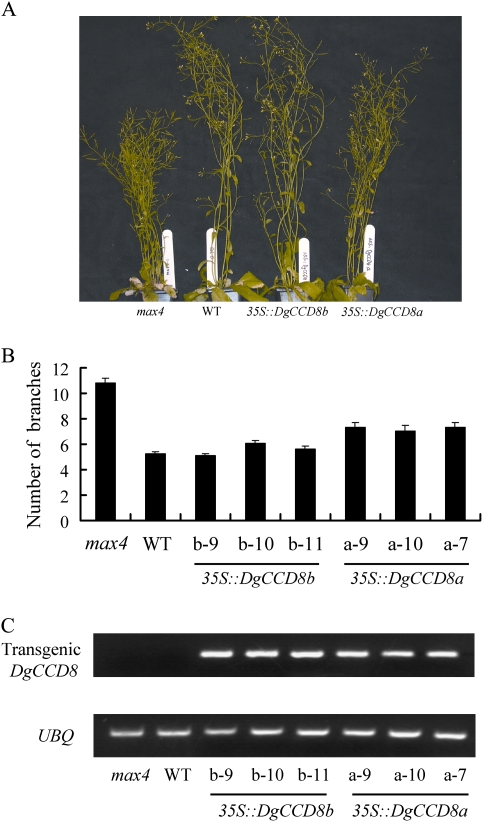
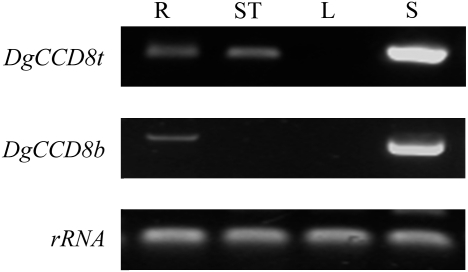
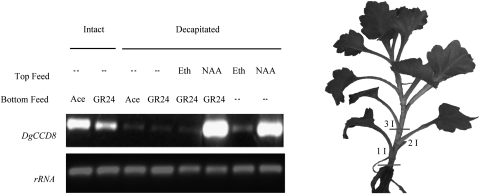
Previous studies of highly branched mutants in pea (rms1-rms5), Arabidopsis thaliana (max1-max4), petunia (dad1-dad3), and rice (d3, d10, htd1/d17, d14, d27) identified strigolactones or their derivates (SLs), as shoot branching inhibitors. This recent discovery offers the possibility of using SLs to regulate branching commercially, for example, in chrysanthemum, an important cut flower crop. To investigate this option, SL physiology and molecular biology were studied in chrysanthemum (Dendranthema grandiflorum), focusing on the CCD8/MAX4/DAD1/RMS1/D10 gene. Our results suggest that, as has been proposed for Arabidopsis, the ability of SLs to inhibit bud activity depends on the presence of a competing auxin source. The chrysanthemum SL biosynthesis gene, CCD8 was cloned, and found to be regulated in a similar, but not identical way to known CCD8s. Expression analyses revealed that DgCCD8 is predominantly expressed in roots and stems, and is up-regulated by exogenous auxin. Exogenous SL can down-regulate DgCCD8 expression, but this effect can be overridden by apical auxin application. This study provides evidence that SLs are promising candidates to alter the shoot branching habit of chrysanthemum.
Figures
References
-
- Alder A, Holdermann I, Beyer P, Al-Babili S. Carotenoid oxygenases involved in plant branching catalyse a highly specific conserved apocarotenoid cleavage reaction. Biochemical Journal. 2008;416:289–296. - PubMed
-
- Arite T, Iwata H, Ohshima K, Maekawa M, Nakajima M, Kojima M, Sakakibara H, Kyozuka J. DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth in rice. The Plant Journal. 2007;51:1019–1029. - PubMed
-
- Arite T, Umehara M, Ishikawa S, Hanada A, Maekawa M, Yamaguchi S, Kyozuka J. d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant and Cell Physiology. 2009;50:1416–1424. - PubMed
-
- Auldridge ME, Block A, Vogel JT, Dabney-Smith C, Mila I, Bouzayen M, Magallanes-Lundback M, DellaPenna D, McCarty DR, Klee HJ. Characterization of three members of the Arabidopsis carotenoid cleavage dioxygenase family demonstrates the divergent roles of this multifunctional enzyme family. The Plant Journal. 2006;45:982–993. - PubMed
-
- Bainbridge K, Sorefan K, Ward S, Leyser O. Hormonally controlled expression of the Arabidopsis MAX4 shoot branching regulatory gene. The Plant Journal. 2005;44:569–580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
