

Nrf2 establishes a glutathione-mediated gradient of UVB cytoprotection in the epidermis
- PMID: 20478997
- PMCID: PMC2867209
- DOI: 10.1101/gad.568810
Nrf2 establishes a glutathione-mediated gradient of UVB cytoprotection in the epidermis
Abstract
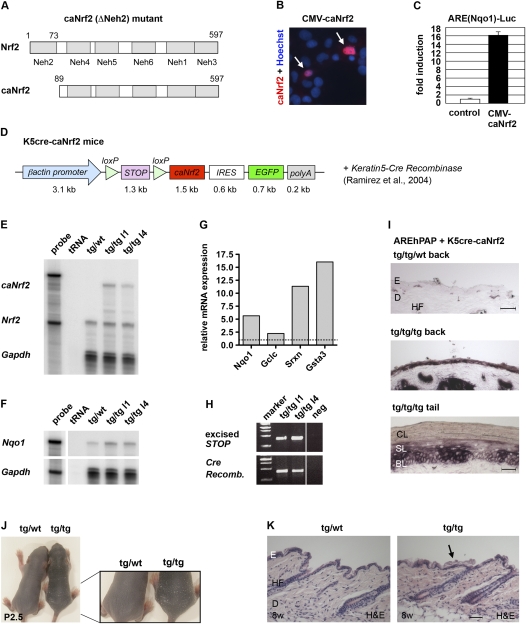
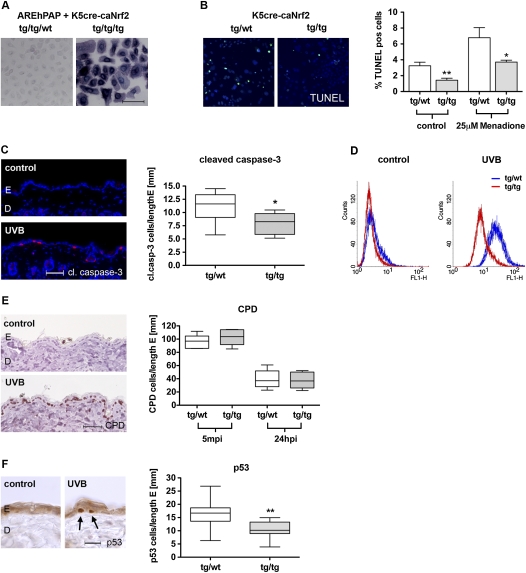
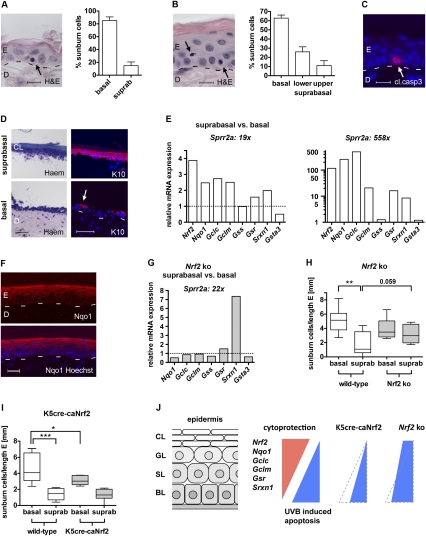
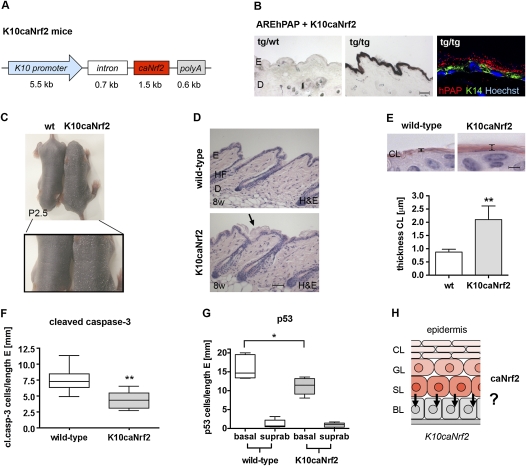
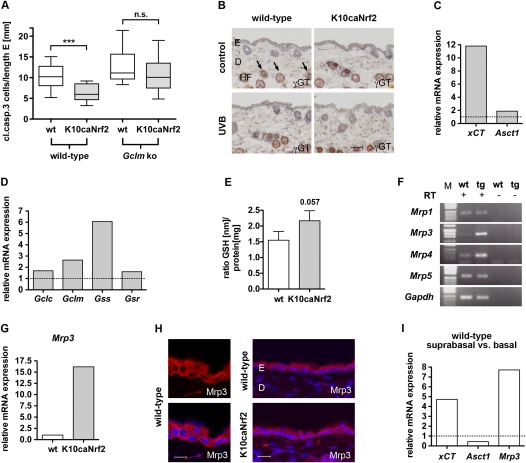
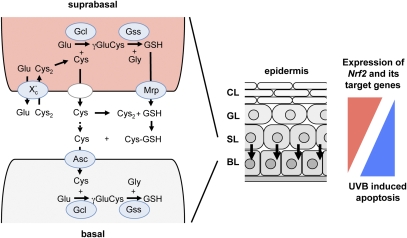
Ultraviolet (UV) B irradiation can severely damage the skin and even induce tumorigenesis. It exerts its effects by direct DNA modification and by formation of reactive oxygen species (ROS). We developed a strategy to genetically activate target gene expression of the transcription factor NF-E2-related factor 2 (Nrf2) in keratinocytes in vivo based on expression of a constitutively active Nrf2 mutant. Activation of Nrf2 target genes strongly reduced UVB cytotoxicity through enhancement of ROS detoxification. Remarkably, the protective effect was extended to neighboring cells. Using different combinations of genetically modified mice, we demonstrate that Nrf2 activates the production, recycling, and release of glutathione and cysteine by suprabasal keratinocytes, resulting in protection of basal cells in a paracrine, glutathione/cysteine-dependent manner. Most importantly, we found that endogenous Nrf2 controls selective protection of suprabasal keratinocytes from UVB-induced apoptosis through activation of cytoprotective genes. This finding explains the preferential UVB-induced apoptosis of basal cells, which is important for elimination of mutated stem cells as well as for preservation of skin integrity. Taken together, our results identify Nrf2 as a key regulator in the UV response of the skin.
Figures
Similar articles
-
Electrophilic chemicals but not UV irradiation or reactive oxygen species activate Nrf2 in keratinocytes in vitro and in vivo.J Invest Dermatol. 2007 Mar;127(3):646-53. doi: 10.1038/sj.jid.5700585. Epub 2006 Sep 28. J Invest Dermatol. 2007. PMID: 17008872
-
Zerumbone protects human skin keratinocytes against UVA-irradiated damages through Nrf2 induction.Biochem Pharmacol. 2018 Feb;148:130-146. doi: 10.1016/j.bcp.2017.12.014. Epub 2017 Dec 20. Biochem Pharmacol. 2018. PMID: 29273513
-
Nrf2 Overexpression for the Protective Effect of Skin-Derived Precursors against UV-Induced Damage: Evidence from a Three-Dimensional Skin Model.Oxid Med Cell Longev. 2019 Oct 14;2019:7021428. doi: 10.1155/2019/7021428. eCollection 2019. Oxid Med Cell Longev. 2019. PMID: 31737172 Free PMC article.
-
Effect of ultraviolet radiation on the Nrf2 signaling pathway in skin cells.Int J Radiat Biol. 2021;97(10):1383-1403. doi: 10.1080/09553002.2021.1962566. Epub 2021 Aug 19. Int J Radiat Biol. 2021. PMID: 34338112 Review.
-
NRF2 in dermatological disorders: Pharmacological activation for protection against cutaneous photodamage and photodermatosis.Free Radic Biol Med. 2022 Aug 1;188:262-276. doi: 10.1016/j.freeradbiomed.2022.06.238. Epub 2022 Jun 23. Free Radic Biol Med. 2022. PMID: 35753587 Free PMC article. Review.
Cited by
-
Ultraviolet Radiation-Induced Skin Aging: The Role of DNA Damage and Oxidative Stress in Epidermal Stem Cell Damage Mediated Skin Aging.Stem Cells Int. 2016;2016:7370642. doi: 10.1155/2016/7370642. Epub 2016 Apr 11. Stem Cells Int. 2016. PMID: 27148370 Free PMC article. Review.
-
Lce1 Family Members Are Nrf2-Target Genes that Are Induced to Compensate for the Loss of Loricrin.J Invest Dermatol. 2016 Aug;136(8):1656-1663. doi: 10.1016/j.jid.2016.04.022. Epub 2016 May 7. J Invest Dermatol. 2016. PMID: 27167730 Free PMC article.
-
Arbutin as a Skin Depigmenting Agent with Antimelanogenic and Antioxidant Properties.Antioxidants (Basel). 2021 Jul 15;10(7):1129. doi: 10.3390/antiox10071129. Antioxidants (Basel). 2021. PMID: 34356362 Free PMC article. Review.
-
The Influence of the Gut Microbiota on Host Physiology: In Pursuit of Mechanisms.Yale J Biol Med. 2016 Sep 30;89(3):285-297. eCollection 2016 Sep. Yale J Biol Med. 2016. PMID: 27698613 Free PMC article. Review.
-
The KEAP1/NRF2 Signaling Pathway in Keratinization.Antioxidants (Basel). 2020 Aug 14;9(8):751. doi: 10.3390/antiox9080751. Antioxidants (Basel). 2020. PMID: 32823937 Free PMC article. Review.
References
-
- Agar N, Young AR 2005. Melanogenesis: A photoprotective response to DNA damage? Mutat Res 571: 121–132 - PubMed
-
- Anderson RR, Parrish JA 1981. The optics of human skin. J Invest Dermatol 77: 13–19 - PubMed
-
- Beyer TA, Auf dem Keller U, Braun S, Schafer M, Werner S 2007. Roles and mechanisms of action of the Nrf2 transcription factor in skin morphogenesis, wound repair and skin cancer. Cell Death Differ 14: 1250–1254 - PubMed
-
- Bickers DR, Athar M 2006. Oxidative stress in the pathogenesis of skin disease. J Invest Dermatol 126: 2565–2575 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases