

Activation of swelling-activated chloride current by tumor necrosis factor-alpha requires ClC-3-dependent endosomal reactive oxygen production
- PMID: 20479003
- PMCID: PMC2906278
- DOI: 10.1074/jbc.M109.099838
Activation of swelling-activated chloride current by tumor necrosis factor-alpha requires ClC-3-dependent endosomal reactive oxygen production
Abstract
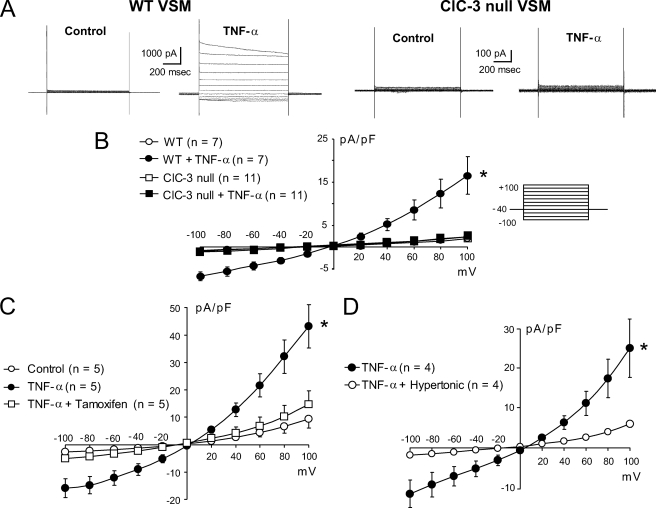
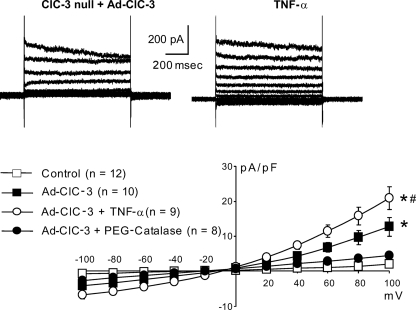
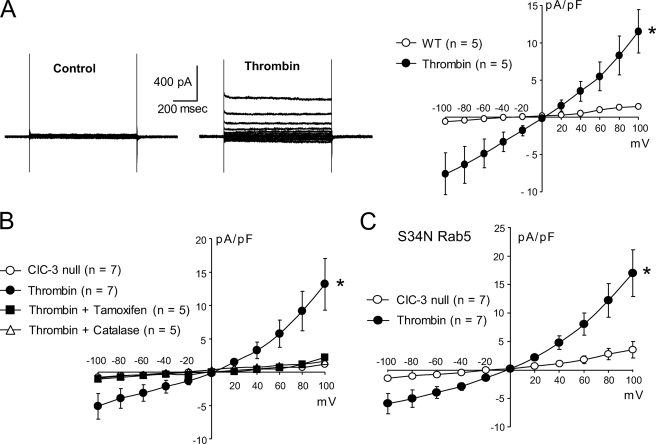
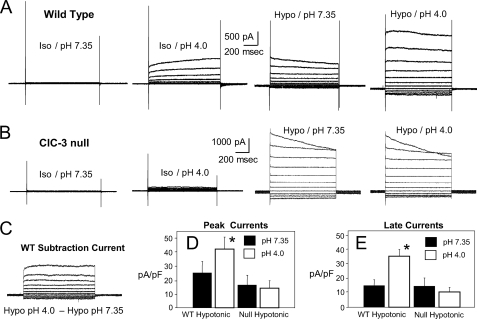
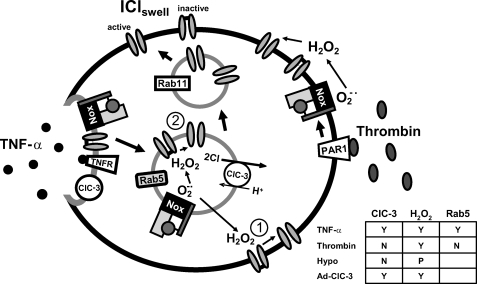
ClC-3 is a Cl(-)/H(+) antiporter required for cytokine-induced intraendosomal reactive oxygen species (ROS) generation by Nox1. ClC-3 current is distinct from the swelling-activated chloride current (ICl(swell)), but overexpression of ClC-3 can activate currents that resemble ICl(swell). Because H(2)O(2) activates ICl(swell) directly, we hypothesized that ClC-3-dependent, endosomal ROS production activates ICl(swell). Whole-cell perforated patch clamp methods were used to record Cl(-) currents in cultured aortic vascular smooth muscle cells from wild type (WT) and ClC-3 null mice. Under isotonic conditions, tumor necrosis factor-alpha (TNF-alpha) (10 ng/ml) activated outwardly rectifying Cl(-) currents with time-dependent inactivation in WT but not ClC-3 null cells. Inhibition by tamoxifen (10 microm) and by hypertonicity (340 mosm) identified them as ICl(swell). ICl(swell) was also activated by H(2)O(2) (500 microm), and the effect of TNF-alpha was completely inhibited by polyethylene glycol-catalase. ClC-3 expression induced ICl(swell) in ClC-3 null cells in the absence of swelling or TNF-alpha, and this effect was also blocked by catalase. ICl(swell) activation by hypotonicity (240 mosm) was only partially inhibited by catalase, and the size of these currents did not differ between WT and ClC-3 null cells. Disruption of endosome trafficking with either mutant Rab5 (S34N) or Rab11 (S25N) inhibited TNF-alpha-mediated activation of ICl(swell). Thrombin also activates ROS production by Nox1 but not in endosomes. Thrombin caused H(2)O(2)-dependent activation of ICl(swell), but this effect was not ClC-3- or Rab5-dependent. Thus, activation of ICl(swell) by TNF-alpha requires ClC-3-dependent endosomal H(2)O(2) production. This demonstrates a functional link between two distinct anion currents, ClC-3 and ICl(swell).
Figures




Similar articles
-
Swelling-activated Cl- currents and intracellular CLC-3 are involved in proliferation of human pulmonary artery smooth muscle cells.J Hypertens. 2014 Feb;32(2):318-30. doi: 10.1097/HJH.0000000000000013. J Hypertens. 2014. PMID: 24284495
-
Inhibition of swelling-activated Cl- currents by functional anti-ClC-3 antibody in native bovine non-pigmented ciliary epithelial cells.Invest Ophthalmol Vis Sci. 2005 Mar;46(3):948-55. doi: 10.1167/iovs.04-1004. Invest Ophthalmol Vis Sci. 2005. PMID: 15728552
-
The role of swelling-activated chloride currents (I(CL,swell)) in the regulatory volume decrease response of freshly dissociated rat articular chondrocytes.Cell Physiol Biochem. 2012;30(5):1254-70. doi: 10.1159/000343316. Epub 2012 Oct 19. Cell Physiol Biochem. 2012. PMID: 23075704
-
The ClC-3 Cl-/H+ antiporter becomes uncoupled at low extracellular pH.J Biol Chem. 2010 Jan 22;285(4):2569-79. doi: 10.1074/jbc.M109.018002. Epub 2009 Nov 19. J Biol Chem. 2010. PMID: 19926787 Free PMC article.
-
Swelling-activated chloride channels in cardiac physiology and pathophysiology.Prog Biophys Mol Biol. 2003 May-Jul;82(1-3):25-42. doi: 10.1016/s0079-6107(03)00003-8. Prog Biophys Mol Biol. 2003. PMID: 12732266 Review.
Cited by
-
The basic biology of redoxosomes in cytokine-mediated signal transduction and implications for disease-specific therapies.Biochemistry. 2014 Mar 18;53(10):1551-64. doi: 10.1021/bi401719r. Epub 2014 Mar 4. Biochemistry. 2014. PMID: 24555469 Free PMC article. Review.
-
Nox1 NADPH oxidase is necessary for late but not early myocardial ischaemic preconditioning.Cardiovasc Res. 2014 Apr 1;102(1):79-87. doi: 10.1093/cvr/cvu027. Epub 2014 Feb 5. Cardiovasc Res. 2014. PMID: 24501329 Free PMC article.
-
LRRC8A channels support TNFα-induced superoxide production by Nox1 which is required for receptor endocytosis.Free Radic Biol Med. 2016 Dec;101:413-423. doi: 10.1016/j.freeradbiomed.2016.11.003. Epub 2016 Nov 9. Free Radic Biol Med. 2016. PMID: 27838438 Free PMC article.
-
Recent insights on the impact of SWELL1 on metabolic syndromes.Front Pharmacol. 2025 Mar 21;16:1552176. doi: 10.3389/fphar.2025.1552176. eCollection 2025. Front Pharmacol. 2025. PMID: 40191429 Free PMC article. Review.
-
Endotoxin priming of neutrophils requires endocytosis and NADPH oxidase-dependent endosomal reactive oxygen species.J Biol Chem. 2012 Apr 6;287(15):12395-404. doi: 10.1074/jbc.M111.306530. Epub 2012 Jan 10. J Biol Chem. 2012. PMID: 22235113 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
