

Ancient origin of the integrin-mediated adhesion and signaling machinery
- PMID: 20479219
- PMCID: PMC2890464
- DOI: 10.1073/pnas.1002257107
Ancient origin of the integrin-mediated adhesion and signaling machinery
Abstract
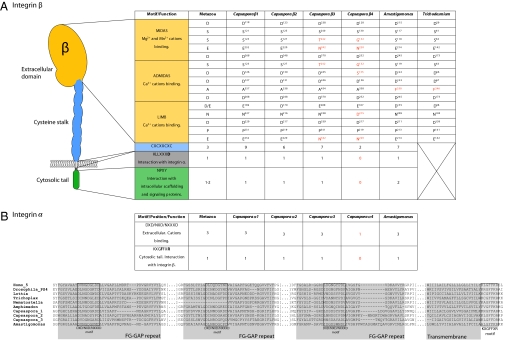
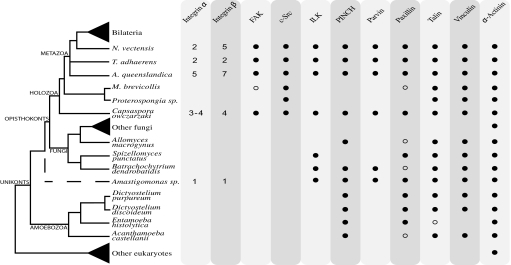
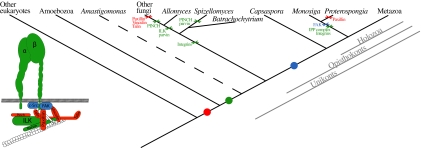
The evolution of animals (metazoans) from their unicellular ancestors required the emergence of novel mechanisms for cell adhesion and cell-cell communication. One of the most important cell adhesion mechanisms for metazoan development is integrin-mediated adhesion and signaling. The integrin adhesion complex mediates critical interactions between cells and the extracellular matrix, modulating several aspects of cell physiology. To date this machinery has been considered strictly metazoan specific. Here we report the results of a comparative genomic analysis of the integrin adhesion machinery, using genomic data from several unicellular relatives of Metazoa and Fungi. Unexpectedly, we found that core components of the integrin adhesion complex are encoded in the genome of the apusozoan protist Amastigomonas sp., and therefore their origins predate the divergence of Opisthokonta, the clade that includes metazoans and fungi. Furthermore, our analyses suggest that key components of this apparatus have been lost independently in fungi and choanoflagellates. Our data highlight the fact that many of the key genes that had formerly been cited as crucial for metazoan origins have a much earlier origin. This underscores the importance of gene cooption in the unicellular-to-multicellular transition that led to the emergence of the Metazoa.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- King N. The unicellular ancestry of animal development. Dev Cell. 2004;7:313–325. - PubMed
-
- Rokas A. The origins of multicellularity and the early history of the genetic toolkit for animal development. Annu Rev Genet. 2008;42:235–251. - PubMed
-
- Putnam NH, et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science. 2007;317:86–94. - PubMed
-
- Srivastava M, et al. The Trichoplax genome and the nature of placozoans. Nature. 2008;454:955–960. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
