

Insights into the recruitment of the NMD machinery from the crystal structure of a core EJC-UPF3b complex
- PMID: 20479275
- PMCID: PMC2890422
- DOI: 10.1073/pnas.1000993107
Insights into the recruitment of the NMD machinery from the crystal structure of a core EJC-UPF3b complex
Abstract
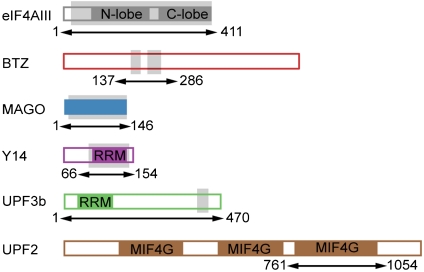
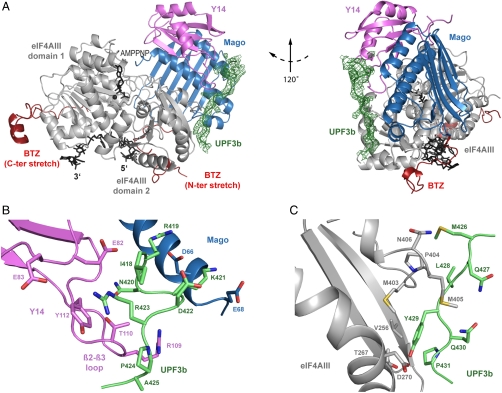
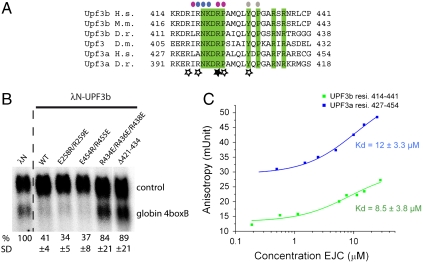
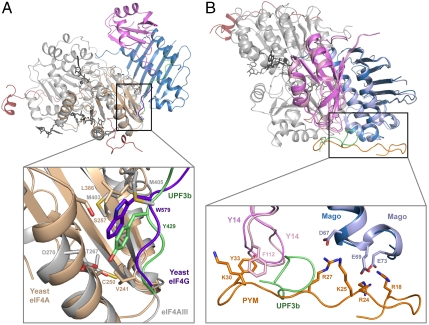
In mammals, Up-frameshift proteins (UPFs) form a surveillance complex that interacts with the exon junction complex (EJC) to elicit nonsense-mediated mRNA decay (NMD). UPF3b is the component of the surveillance complex that bridges the interaction with the EJC. Here, we report the 3.4 A resolution crystal structure of a minimal UPF3b-EJC assembly, consisting of the interacting domains of five proteins (UPF3b, MAGO, Y14, eIF4AIII, and Barentsz) together with RNA and adenylyl-imidodiphosphate. Human UPF3b binds with the C-terminal domain stretched over a composite surface formed by eIF4AIII, MAGO, and Y14. Residues that affect NMD when mutated are found at the core interacting surfaces, whereas differences between UPF3b and UPF3a map at peripheral interacting residues. Comparison with the binding mode of the protein PYM underscores how a common molecular surface of MAGO and Y14 recognizes different proteins acting at different times in the same pathway. The binding mode to eIF4AIII identifies a surface hot spot that is used by different DEAD-box proteins to recruit their regulators.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Mutational analysis of human eIF4AIII identifies regions necessary for exon junction complex formation and nonsense-mediated mRNA decay.RNA. 2006 Mar;12(3):360-74. doi: 10.1261/rna.2190706. RNA. 2006. PMID: 16495234 Free PMC article.
-
The crystal structure of the exon junction complex reveals how it maintains a stable grip on mRNA.Cell. 2006 Aug 25;126(4):713-25. doi: 10.1016/j.cell.2006.08.006. Cell. 2006. PMID: 16923391
-
Functions of hUpf3a and hUpf3b in nonsense-mediated mRNA decay and translation.RNA. 2006 Jun;12(6):1015-22. doi: 10.1261/rna.12506. Epub 2006 Apr 6. RNA. 2006. PMID: 16601204 Free PMC article.
-
New insights into the formation of active nonsense-mediated decay complexes.Trends Biochem Sci. 2003 Sep;28(9):464-6. doi: 10.1016/S0968-0004(03)00176-2. Trends Biochem Sci. 2003. PMID: 13678954 Review.
-
Structural insights into the exon junction complex.Curr Opin Struct Biol. 2008 Feb;18(1):112-9. doi: 10.1016/j.sbi.2007.11.002. Curr Opin Struct Biol. 2008. PMID: 18164611 Review.
Cited by
-
Nonsense-mediated RNA decay: an emerging modulator of malignancy.Nat Rev Cancer. 2022 Aug;22(8):437-451. doi: 10.1038/s41568-022-00481-2. Epub 2022 May 27. Nat Rev Cancer. 2022. PMID: 35624152 Free PMC article. Review.
-
Quality and quantity control of gene expression by nonsense-mediated mRNA decay.Nat Rev Mol Cell Biol. 2019 Jul;20(7):406-420. doi: 10.1038/s41580-019-0126-2. Nat Rev Mol Cell Biol. 2019. PMID: 30992545 Free PMC article. Review.
-
UPF1 helicase orchestrates mutually exclusive interactions with the SMG6 endonuclease and UPF2.Nucleic Acids Res. 2024 Jun 10;52(10):6036-6048. doi: 10.1093/nar/gkae323. Nucleic Acids Res. 2024. PMID: 38709891 Free PMC article.
-
DEAD-box proteins as RNA helicases and chaperones.Wiley Interdiscip Rev RNA. 2011 Jan-Feb;2(1):135-52. doi: 10.1002/wrna.50. Wiley Interdiscip Rev RNA. 2011. PMID: 21297876 Free PMC article. Review.
-
Comparison of EJC-enhanced and EJC-independent NMD in human cells reveals two partially redundant degradation pathways.RNA. 2013 Oct;19(10):1432-48. doi: 10.1261/rna.038893.113. Epub 2013 Aug 20. RNA. 2013. PMID: 23962664 Free PMC article.
References
-
- Chang YF, Imam JS, Wilkinson MF. The nonsense-mediated decay RNA surveillance pathway. Annu Rev Biochem. 2007;76:51–74. - PubMed
-
- Behm-Ansmant I, Izaurralde E. Quality control of gene expression: A stepwise assembly pathway for the surveillance complex that triggers nonsense-mediated mRNA decay. Genes Dev. 2006;20:391–398. - PubMed
-
- Conti E, Izaurralde E. Nonsense-mediated mRNA decay: Molecular insights and mechanistic variations across species. Curr Opin Cell Biol. 2005;17:316–325. - PubMed
-
- Muhlemann O, Eberle AB, Stalder L, Zamudio Orozco R. Recognition and elimination of nonsense mRNA. Biochim Biophys Acta. 2008;1779:538–549. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
