

Straight-chain alkyl isocyanides open the distal histidine gate in crystal structures of myoglobin
- PMID: 20481504
- PMCID: PMC4074459
- DOI: 10.1021/bi1001739
Straight-chain alkyl isocyanides open the distal histidine gate in crystal structures of myoglobin
Abstract
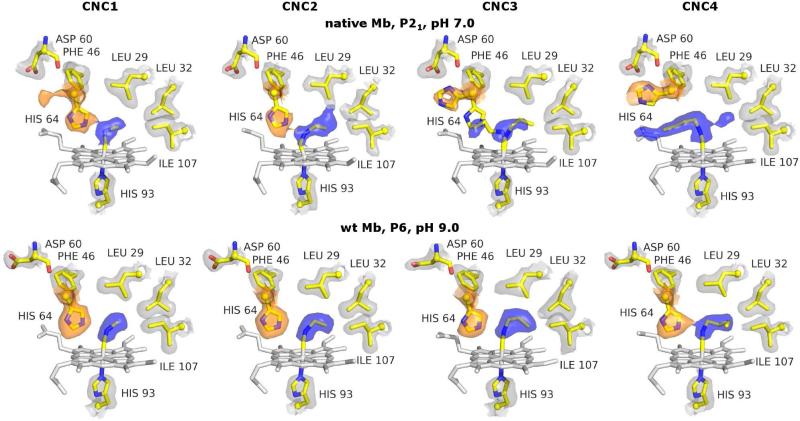
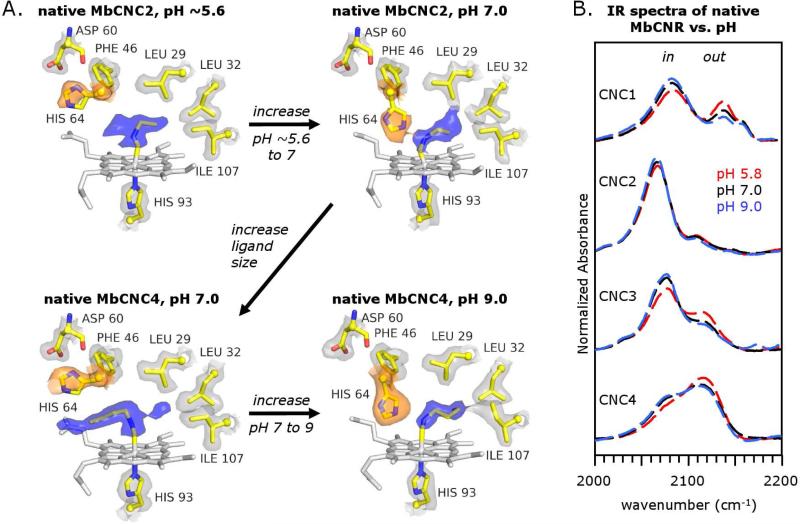
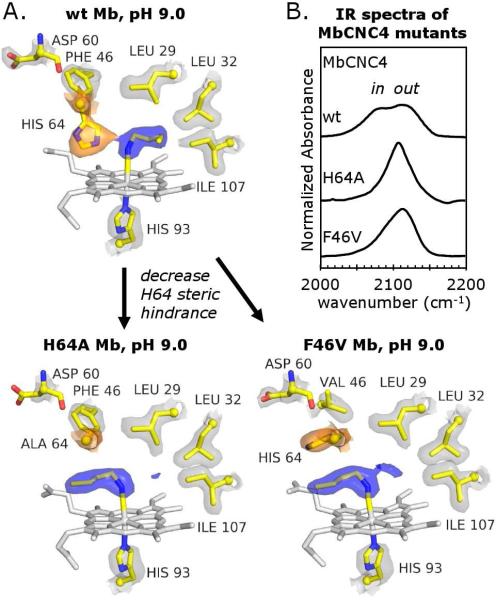
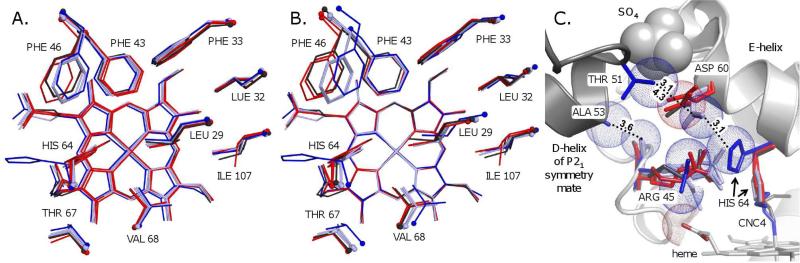
Crystal structures of methyl, ethyl, propyl, and butyl isocyanide bound to sperm whale myoglobin (Mb) reveal two major conformations. In the in conformer, His(E7) is in a "closed" position, forcing the ligand alkyl chain to point inward. In the out conformer, His(E7) is in an "open" position, allowing the ligand side chain to point outward. A progressive increase in the population of the out conformer is observed with increasing ligand length in P2(1) crystals of native Mb at pH 7.0. This switch from in to out with increasing ligand size also occurs in solution as measured by the decrease in the relative intensity of the low-frequency ( approximately 2075 cm(-1)) versus high-frequency ( approximately 2125 cm(-1)) isocyano bands. In contrast, all four isocyanides in P6 crystals of wild-type recombinant Mb occupy the in conformation. However, mutating either His64 to Ala, creating a "hole" to solvent, or Phe46 to Val, freeing rotation of His64, causes bound butyl isocyanide to point completely outward in P6 crystals. Thus, the unfavorable hindrance caused with crowding a large alkyl side chain into the distal pocket appears to be roughly equal to that for pushing open the His(E7) gate and is easily affected by crystal packing. This structural conclusion supports the "side path" kinetic mechanism for O(2) release, in which the dissociated ligand first moves toward the protein interior and then encounters steric resistance, which is roughly equal to that for escaping to solvent through the His(E7) channel.
Figures
References
-
- Dou Y, Maillett DH, Eich RF, Olson JS. Myoglobin as a model system for designing heme protein based blood substitutes. Biophys Chem. 2002;98:127–148. - PubMed
-
- Bellelli A, Brunori M, Miele AE, Panetta G, Vallone B. The allosteric properties of hemoglobin: insights from natural and site directed mutants. Curr Protein Pept Sci. 2006;7:17–45. - PubMed
-
- Hankeln T, Ebner B, Fuchs C, Gerlach F, Haberkamp M, Laufs TL, Roesner A, Schmidt M, Weich B, Wystub S, Saaler-Reinhardt S, Reuss S, Bolognesi M, De Sanctis D, Marden MC, Kiger L, Moens L, Dewilde S, Nevo E, Avivi A, Weber RE, Fago A, Burmester T. Neuroglobin and cytoglobin in search of their role in the vertebrate globin family. J Inorg Biochem. 2005;99:110–119. - PubMed
-
- Hoy JA, Hargrove MS. The structure and function of plant hemoglobins. Plant Physiol Biochem. 2008;46:371–379. - PubMed
-
- Hoy JA, Robinson H, Trent JT, 3rd, Kakar S, Smagghe BJ, Hargrove MS. Plant hemoglobins: a molecular fossil record for the evolution of oxygen transport. J Mol Biol. 2007;371:168–179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
