

DNA translocation and unzipping through a nanopore: some geometrical effects
- PMID: 20483325
- PMCID: PMC2872221
- DOI: 10.1016/j.bpj.2010.01.041
DNA translocation and unzipping through a nanopore: some geometrical effects
Abstract
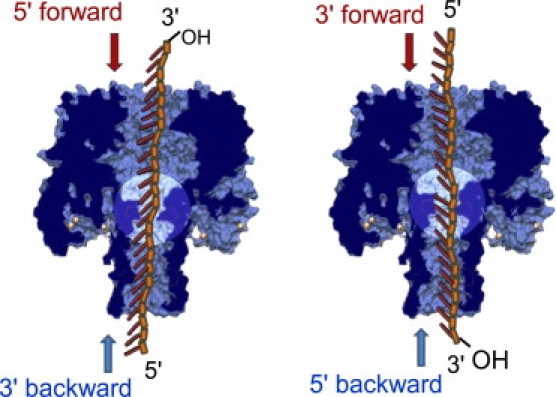
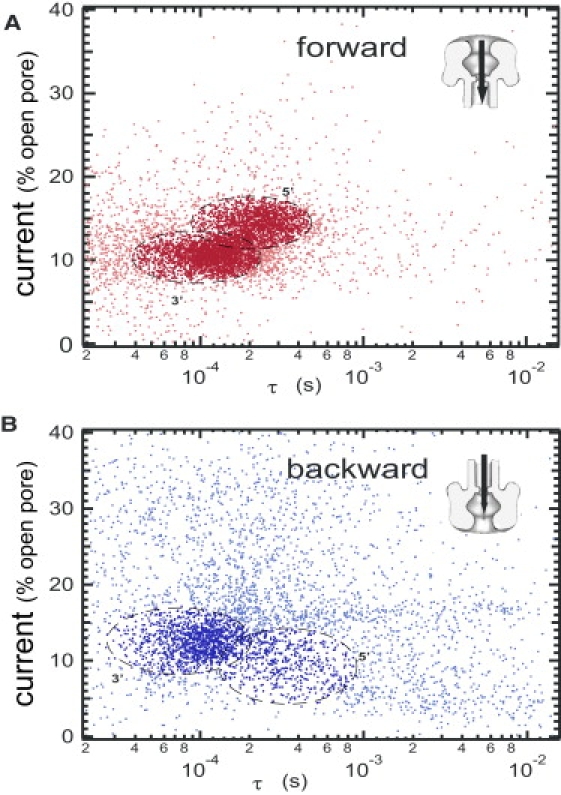
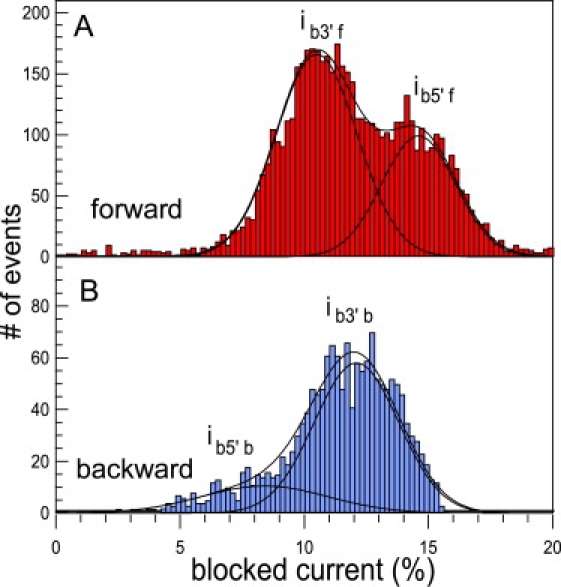
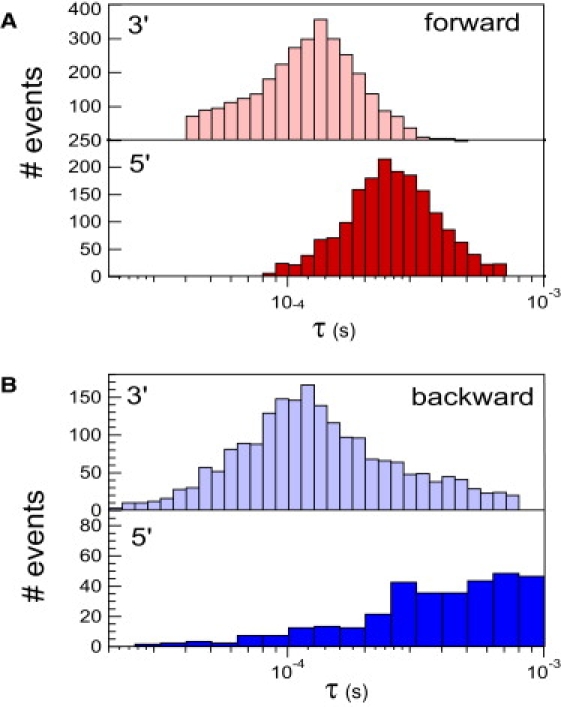
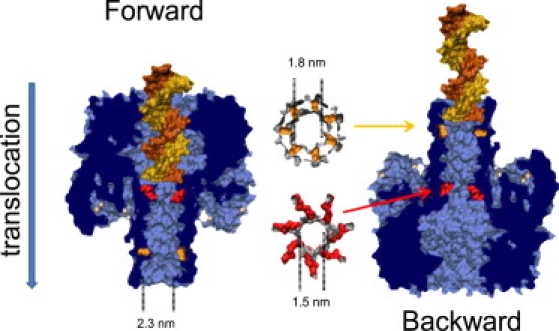
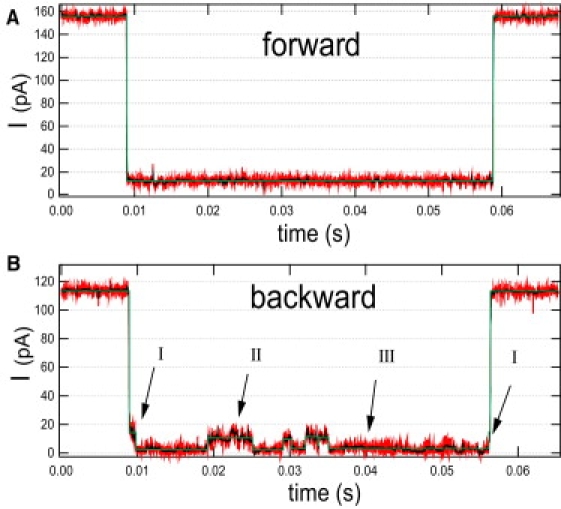
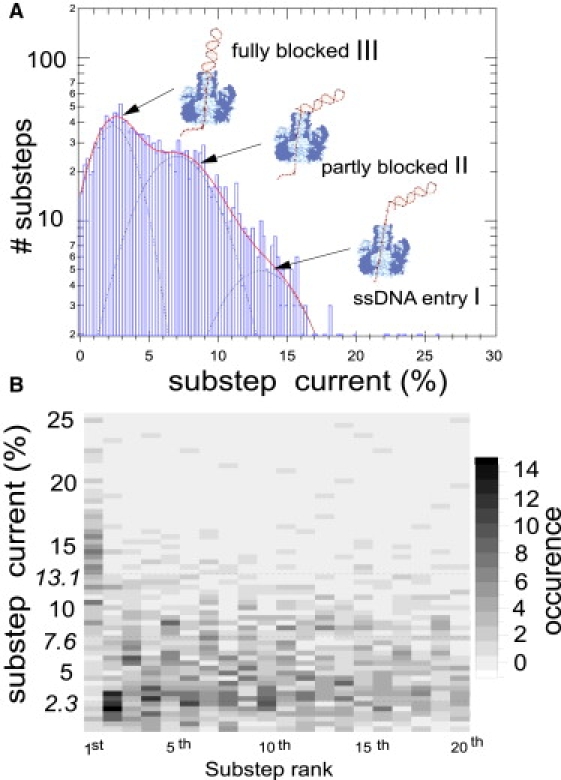
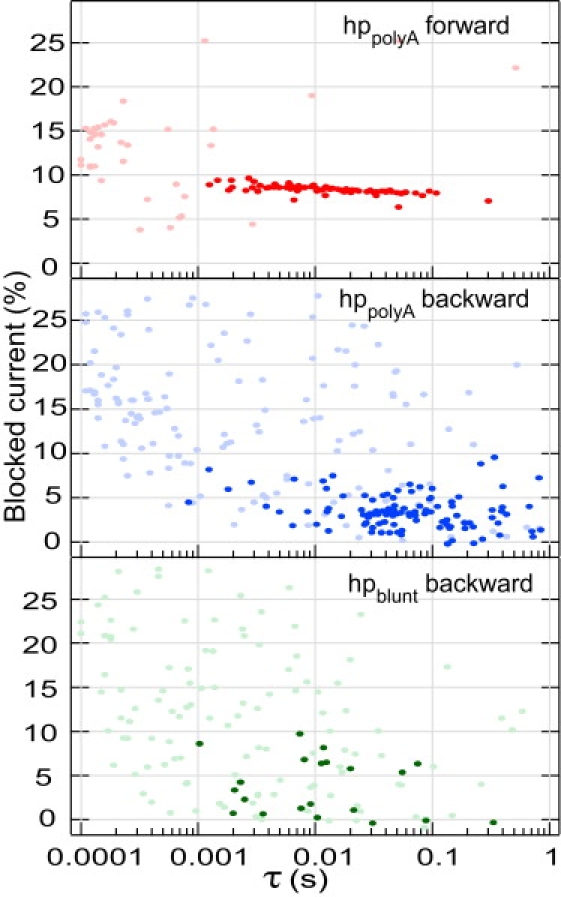
This article explores the role of some geometrical factors on the electrophoretically driven translocations of macromolecules through nanopores. In the case of asymmetric pores, we show how the entry requirements and the direction of translocation can modify the information content of the blocked ionic current as well as the transduction of the electrophoretic drive into a mechanical force. To address these effects we studied the translocation of single-stranded DNA through an asymmetric alpha-hemolysin pore. Depending on the direction of the translocation, we measure the capacity of the pore to discriminate between both DNA orientations. By unzipping DNA hairpins from both sides of the pores we show that the presence of single strand or double strand in the pore can be discriminated based on ionic current levels. We also show that the transduction of the electrophoretic drive into a denaturing mechanical force depends on the local geometry of the pore entrance. Eventually we discuss the application of this work to the measurement of energy barriers for DNA unzipping as well as for protein binding and unfolding.
Copyright 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Internal vs fishhook hairpin DNA: unzipping locations and mechanisms in the α-hemolysin nanopore.J Phys Chem B. 2014 Nov 13;118(45):12873-82. doi: 10.1021/jp5101413. Epub 2014 Nov 3. J Phys Chem B. 2014. PMID: 25333648 Free PMC article.
-
Microscopic Kinetics of DNA Translocation through synthetic nanopores.Biophys J. 2004 Sep;87(3):2086-97. doi: 10.1529/biophysj.104.042960. Biophys J. 2004. PMID: 15345583 Free PMC article.
-
Electrokinetic-flow-induced viscous drag on a tethered DNA inside a nanopore.Phys Rev E Stat Nonlin Soft Matter Phys. 2007 Dec;76(6 Pt 1):061916. doi: 10.1103/PhysRevE.76.061916. Epub 2007 Dec 26. Phys Rev E Stat Nonlin Soft Matter Phys. 2007. PMID: 18233878
-
DNA unzipping and protein unfolding using nanopores.Methods Mol Biol. 2012;870:55-75. doi: 10.1007/978-1-61779-773-6_4. Methods Mol Biol. 2012. PMID: 22528258 Review.
-
Nanopore sequencing technology: nanopore preparations.Trends Biotechnol. 2007 Apr;25(4):174-81. doi: 10.1016/j.tibtech.2007.02.008. Epub 2007 Feb 22. Trends Biotechnol. 2007. PMID: 17320228 Review.
Cited by
-
Kinetics of T3-DNA Ligase-Catalyzed Phosphodiester Bond Formation Measured Using the α-Hemolysin Nanopore.ACS Nano. 2016 Dec 27;10(12):11127-11135. doi: 10.1021/acsnano.6b05995. Epub 2016 Dec 2. ACS Nano. 2016. PMID: 28024377 Free PMC article.
-
Quantitative analysis of the nanopore translocation dynamics of simple structured polynucleotides.Biophys J. 2012 Jan 4;102(1):85-95. doi: 10.1016/j.bpj.2011.11.4011. Epub 2012 Jan 3. Biophys J. 2012. PMID: 22225801 Free PMC article.
-
Temperature and electrolyte optimization of the α-hemolysin latch sensing zone for detection of base modification in double-stranded DNA.Biophys J. 2014 Aug 19;107(4):924-31. doi: 10.1016/j.bpj.2014.07.006. Biophys J. 2014. PMID: 25140427 Free PMC article.
-
Crown ether-electrolyte interactions permit nanopore detection of individual DNA abasic sites in single molecules.Proc Natl Acad Sci U S A. 2012 Jul 17;109(29):11504-9. doi: 10.1073/pnas.1201669109. Epub 2012 Jun 18. Proc Natl Acad Sci U S A. 2012. PMID: 22711805 Free PMC article.
-
On the Lubensky-Nelson model of polymer translocation through nanopores.Biophys J. 2012 Sep 5;103(5):889-97. doi: 10.1016/j.bpj.2012.07.036. Biophys J. 2012. PMID: 23009838 Free PMC article.
References
-
- Stefureac R., Long Y.T., Lee J.S. Transport of α-helical peptides through α-hemolysin and aerolysin pores. Biochemistry. 2006;45:9172–9179. - PubMed
-
- Oukhaled G., Mathé J., Auvray L. Unfolding of proteins and long transient conformations detected by single nanopore recording. Phys. Rev. Lett. 2007;98:158101–158105. - PubMed
-
- Brun L., Pastoriza-Gallego M., Pelta J. Dynamics of polyelectrolyte transport through a protein channel as a function of applied voltage. Phys. Rev. Lett. 2008;100:158302. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
