

ADF/cofilin binds phosphoinositides in a multivalent manner to act as a PIP(2)-density sensor
- PMID: 20483342
- PMCID: PMC2872272
- DOI: 10.1016/j.bpj.2010.01.046
ADF/cofilin binds phosphoinositides in a multivalent manner to act as a PIP(2)-density sensor
Abstract
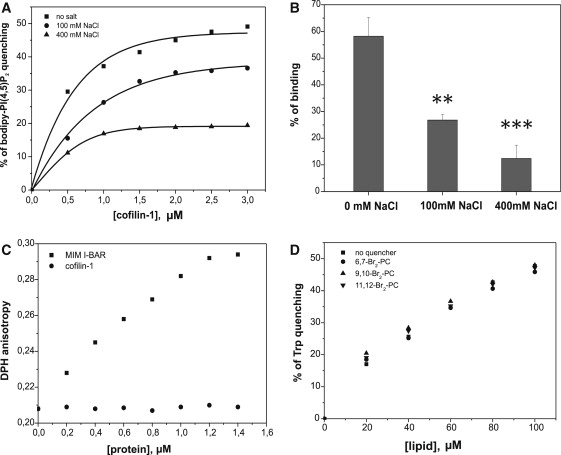
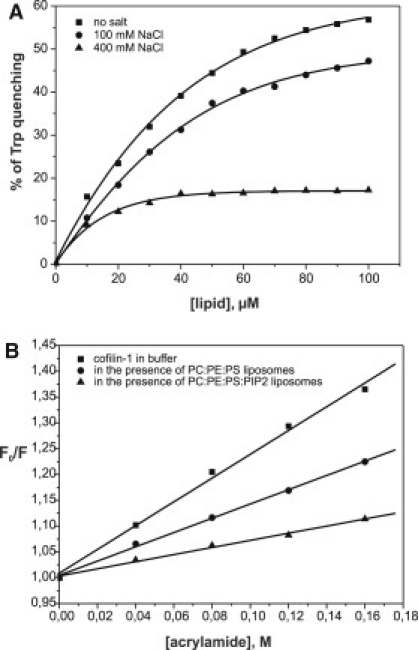
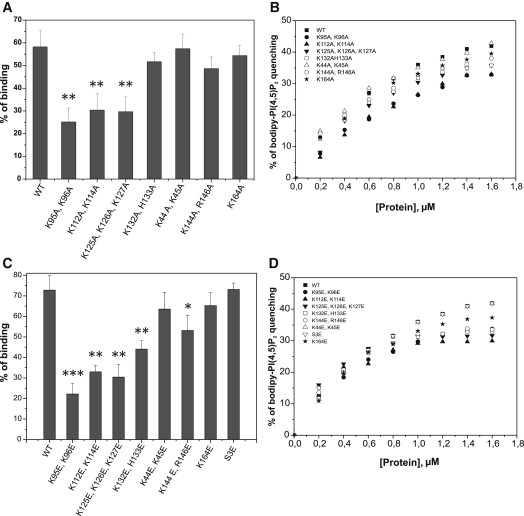
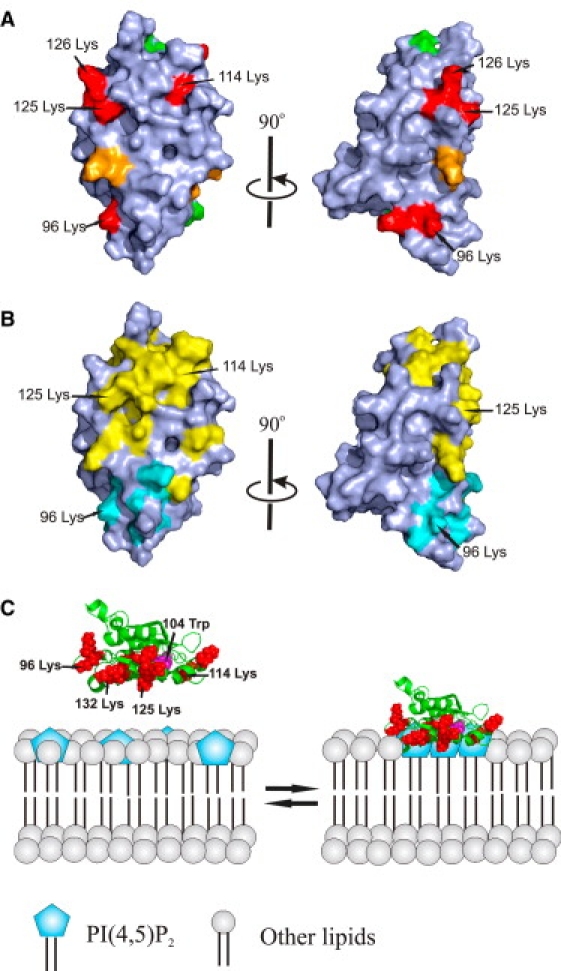
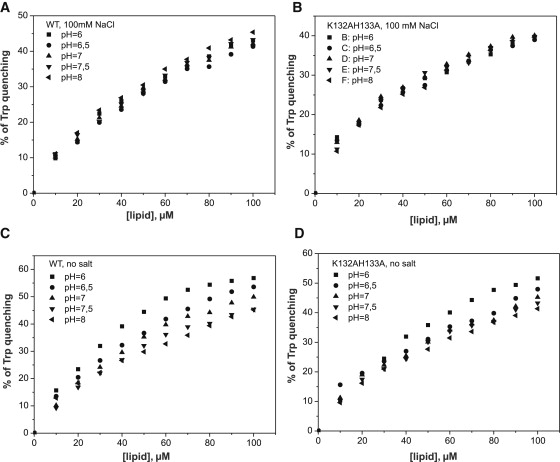
Actin-depolymerizing-factor (ADF)/cofilins have emerged as key regulators of cytoskeletal dynamics in cell motility, morphogenesis, endocytosis, and cytokinesis. The activities of ADF/cofilins are regulated by membrane phospholipid PI(4,5)P(2) in vitro and in cells, but the mechanism of the ADF/cofilin-PI(4,5)P(2) interaction has remained controversial. Recent studies suggested that ADF/cofilins interact with PI(4,5)P(2) through a specific binding pocket, and that this interaction is dependent on pH. Here, we combined systematic mutagenesis with biochemical and spectroscopic methods to elucidate the phosphoinositide-binding mechanism of ADF/cofilins. Our analysis revealed that cofilin does not harbor a specific PI(4,5)P(2)-binding pocket, but instead interacts with PI(4,5)P(2) through a large, positively charged surface of the molecule. Cofilin interacts simultaneously with multiple PI(4,5)P(2) headgroups in a cooperative manner. Consequently, interactions of cofilin with membranes and actin exhibit sharp sensitivity to PI(4,5)P(2) density. Finally, we show that cofilin binding to PI(4,5)P(2) is not sensitive to changes in the pH at physiological salt concentration, although the PI(4,5)P(2)-clustering activity of cofilin is moderately inhibited at elevated pH. Collectively, our data demonstrate that ADF/cofilins bind PI(4,5)P(2) headgroups through a multivalent, cooperative mechanism, and suggest that the actin filament disassembly activity of ADF/cofilin can be accurately regulated by small changes in the PI(4,5)P(2) density at cellular membranes.
Copyright 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



Similar articles
-
The three mouse actin-depolymerizing factor/cofilins evolved to fulfill cell-type-specific requirements for actin dynamics.Mol Biol Cell. 2002 Jan;13(1):183-94. doi: 10.1091/mbc.01-07-0331. Mol Biol Cell. 2002. PMID: 11809832 Free PMC article.
-
Identification of yeast cofilin residues specific for actin monomer and PIP2 binding.Biochemistry. 2001 Dec 25;40(51):15562-9. doi: 10.1021/bi0117697. Biochemistry. 2001. PMID: 11747431
-
Solution structure of human cofilin: actin binding, pH sensitivity, and relationship to actin-depolymerizing factor.J Biol Chem. 2004 Feb 6;279(6):4840-8. doi: 10.1074/jbc.M310148200. Epub 2003 Nov 18. J Biol Chem. 2004. PMID: 14627701
-
Proteins of the ADF/cofilin family: essential regulators of actin dynamics.Annu Rev Cell Dev Biol. 1999;15:185-230. doi: 10.1146/annurev.cellbio.15.1.185. Annu Rev Cell Dev Biol. 1999. PMID: 10611961 Review.
-
The ADF/cofilin family: actin-remodeling proteins.Genome Biol. 2002;3(5):reviews3007. doi: 10.1186/gb-2002-3-5-reviews3007. Epub 2002 Apr 26. Genome Biol. 2002. PMID: 12049672 Free PMC article. Review.
Cited by
-
Incorporation of cofilin into rods depends on disulfide intermolecular bonds: implications for actin regulation and neurodegenerative disease.J Neurosci. 2012 May 9;32(19):6670-81. doi: 10.1523/JNEUROSCI.6020-11.2012. J Neurosci. 2012. PMID: 22573689 Free PMC article.
-
PIP5K-Ras bistability initiates plasma membrane symmetry breaking to regulate cell polarity and migration.bioRxiv [Preprint]. 2024 Sep 15:2024.09.15.613115. doi: 10.1101/2024.09.15.613115. bioRxiv. 2024. PMID: 39314378 Free PMC article. Preprint.
-
The initiation of post-synaptic protrusions.Commun Integr Biol. 2016 Apr 12;9(3):e1125053. doi: 10.1080/19420889.2015.1125053. eCollection 2016 May-Jun. Commun Integr Biol. 2016. PMID: 27489575 Free PMC article.
-
Actin-interacting Protein 1 Promotes Disassembly of Actin-depolymerizing Factor/Cofilin-bound Actin Filaments in a pH-dependent Manner.J Biol Chem. 2016 Mar 4;291(10):5146-56. doi: 10.1074/jbc.M115.713495. Epub 2016 Jan 8. J Biol Chem. 2016. PMID: 26747606 Free PMC article.
-
The Role of ADF/Cofilin in Synaptic Physiology and Alzheimer's Disease.Front Cell Dev Biol. 2020 Nov 12;8:594998. doi: 10.3389/fcell.2020.594998. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33282872 Free PMC article. Review.
References
-
- Bamburg J.R., Wiggan O.P. ADF/cofilin and actin dynamics in disease. Trends Cell Biol. 2002;12:598–605. - PubMed
-
- Friedl P., Gilmour D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2009;10:445–457. - PubMed
-
- Pollard T.D. The cytoskeleton, cellular motility and the reductionist agenda. Nature. 2003;422:741–745. - PubMed
-
- Paavilainen V.O., Bertling E., Lappalainen P. Regulation of cytoskeletal dynamics by actin-monomer-binding proteins. Trends Cell Biol. 2004;14:386–394. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
