

Phagocytosis and phagosome acidification are required for pathogen processing and MyD88-dependent responses to Staphylococcus aureus
- PMID: 20483752
- PMCID: PMC2935932
- DOI: 10.4049/jimmunol.1000110
Phagocytosis and phagosome acidification are required for pathogen processing and MyD88-dependent responses to Staphylococcus aureus
Abstract
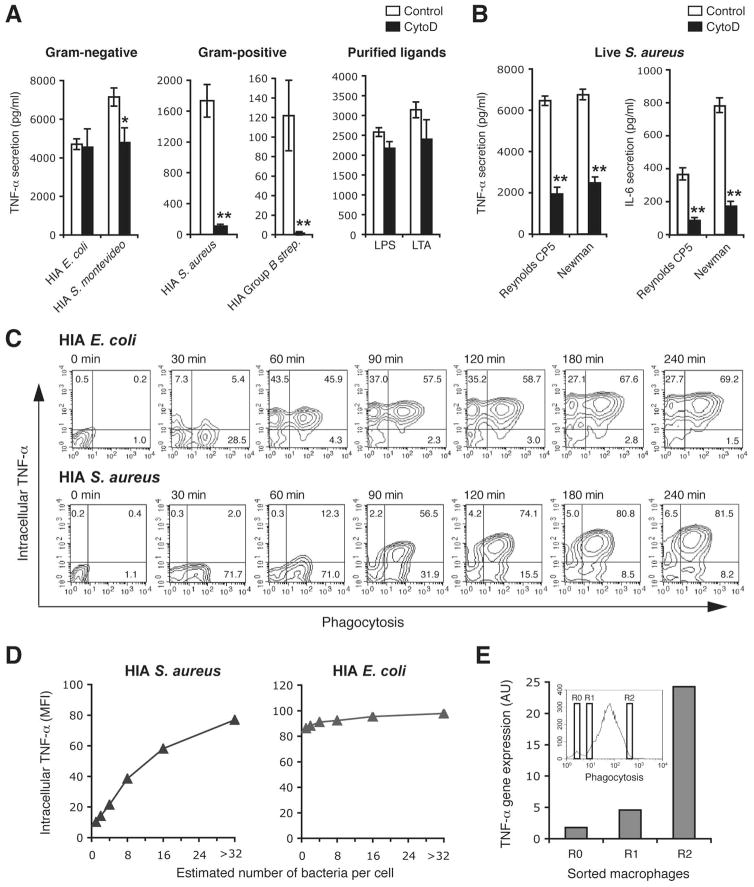
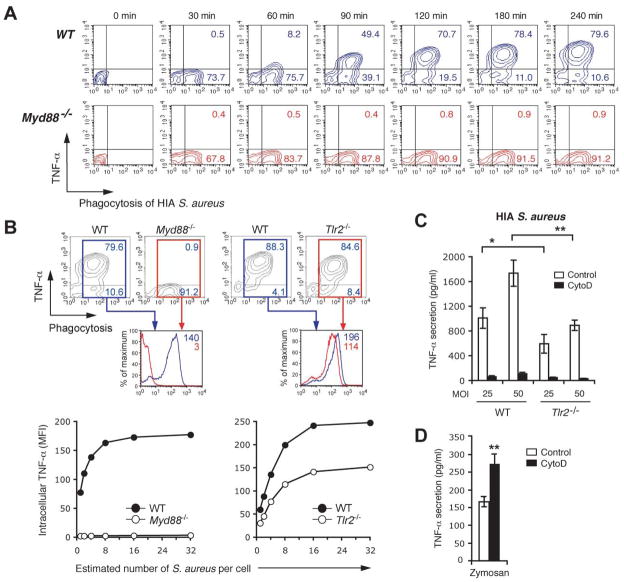
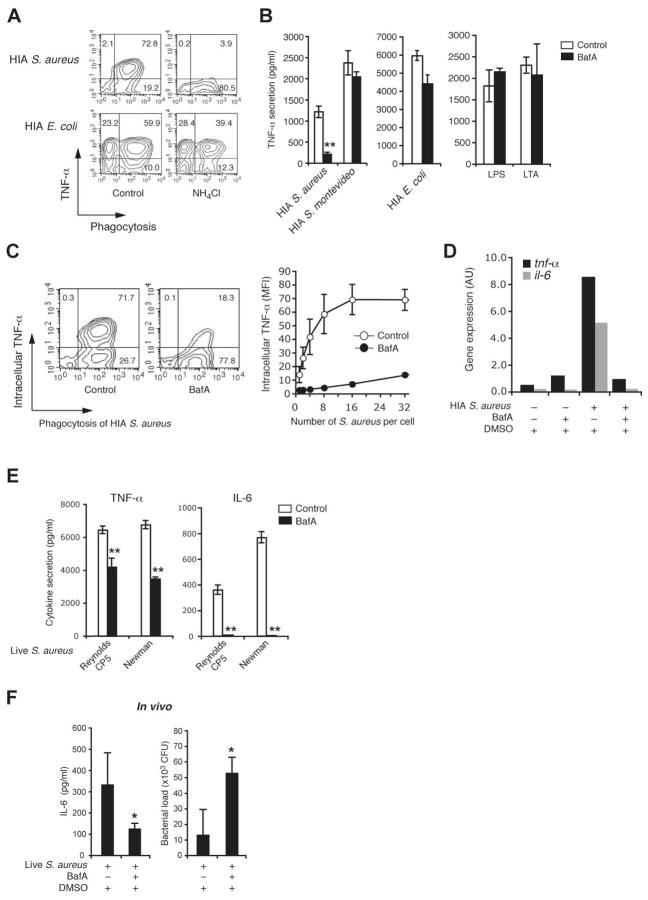
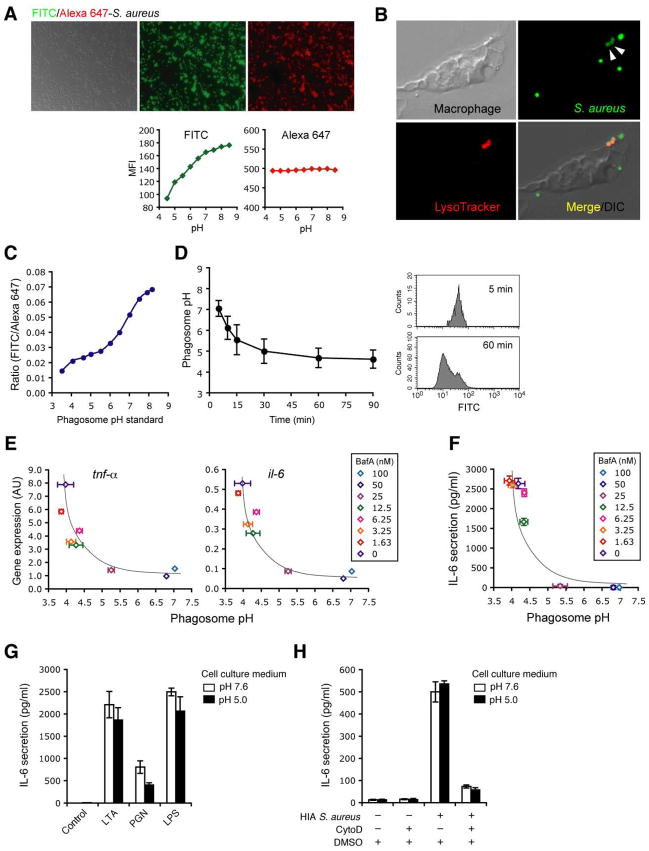
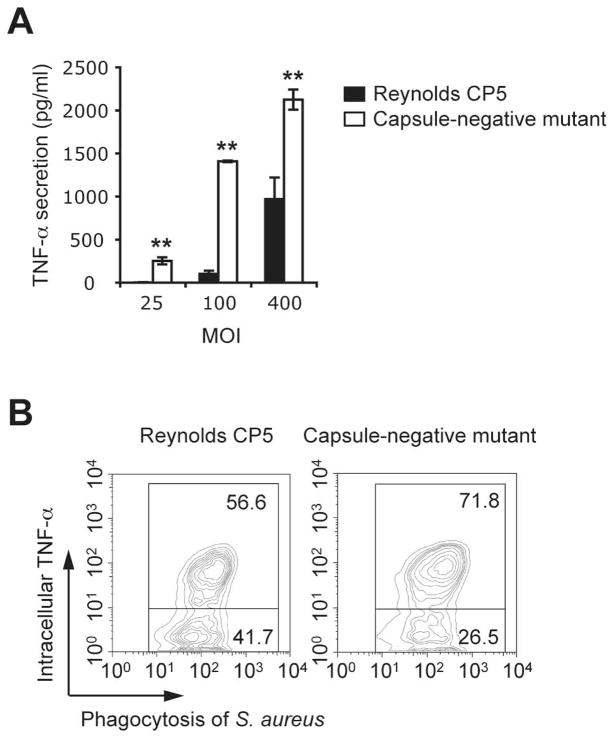
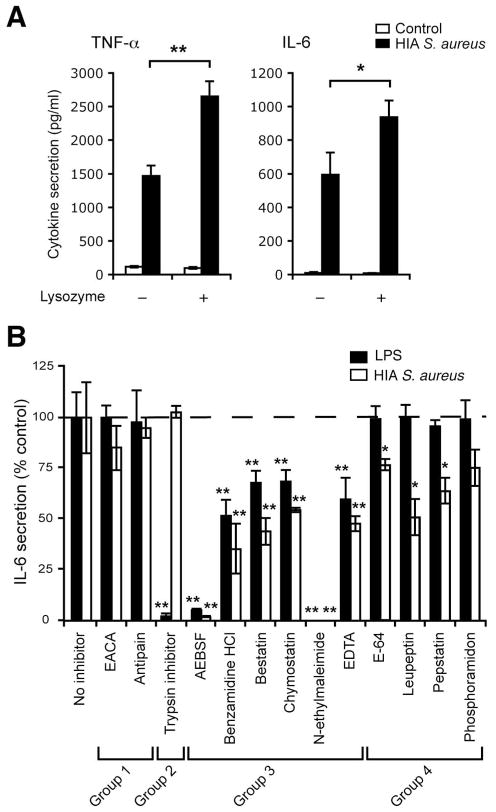
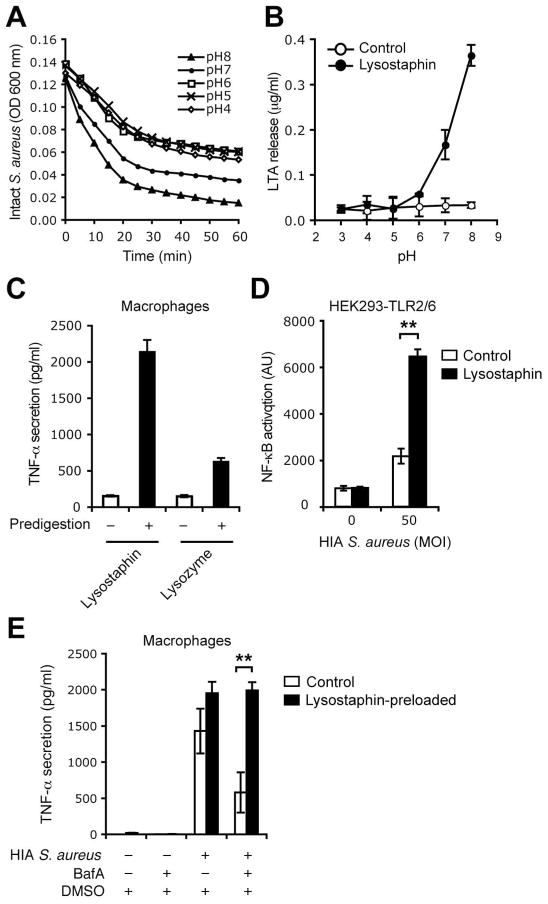
Innate immunity is vital for protection from microbes and is mediated by humoral effectors, such as cytokines, and cellular immune defenses, including phagocytic cells (e.g., macrophages). After internalization by phagocytes, microbes are delivered into a phagosome, a complex intracellular organelle with a well-established and important role in microbial killing. However, the role of this organelle in cytokine responses and microbial sensing is less well defined. In this study, we assess the role of the phagosome in innate immune sensing and demonstrate the critical interdependence of phagocytosis and pattern recognition receptor signaling during response to the Gram-positive bacteria Staphylococcus aureus. We show that phagocytosis is essential to initiate an optimal MyD88-dependent response to Staphylococcus aureus. Prior to TLR-dependent cytokine production, bacteria must be engulfed and delivered into acidic phagosomes where acid-activated host enzymes digest the internalized bacteria to liberate otherwise cryptic bacterial-derived ligands that initiate responses from the vacuole. Importantly, in macrophages in which phagosome acidification is perturbed, the impaired response to S. aureus can be rescued by the addition of lysostaphin, a bacterial endopeptidase active at neutral pH that can substitute for the acid-activated host enzymes. Together, these observations delineate the interdependence of phagocytosis with pattern recognition receptor signaling and suggest that therapeutics to augment functions and signaling from the vacuole may be useful strategies to increase host responses to S. aureus.
Figures
References
-
- Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593–623. - PubMed
-
- Greenberg S, Grinstein S. Phagocytosis and innate immunity. Curr Opin Immunol. 2002;14:1366–145. - PubMed
-
- Stuart LM, Ezekowitz RA. Phagocytosis: elegant complexity. Immunity. 2005;22:539–550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
