

The prostaglandin E2 E-prostanoid 4 receptor exerts anti-inflammatory effects in brain innate immunity
- PMID: 20483760
- PMCID: PMC3103215
- DOI: 10.4049/jimmunol.0903487
The prostaglandin E2 E-prostanoid 4 receptor exerts anti-inflammatory effects in brain innate immunity
Abstract
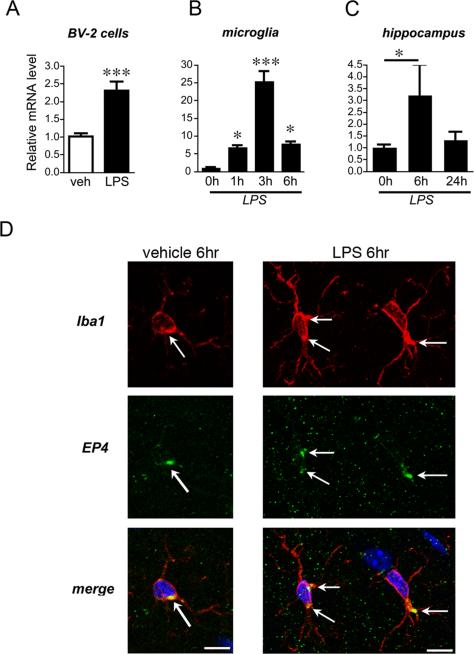
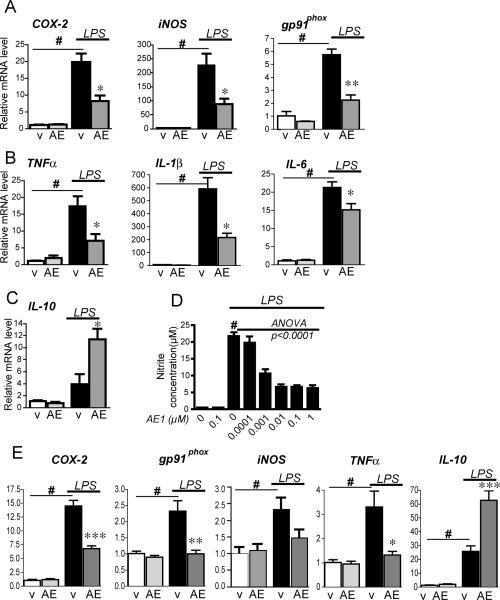
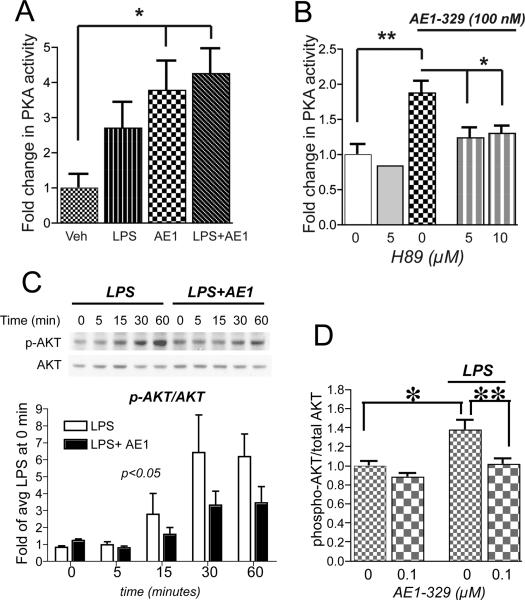
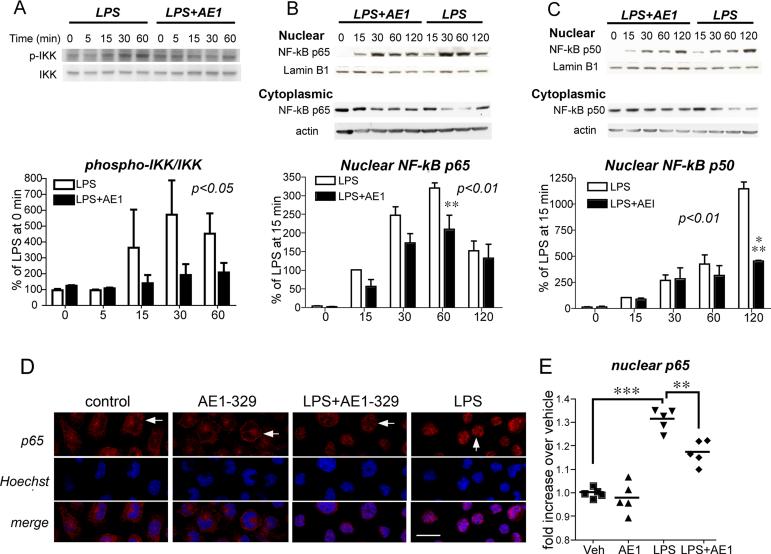
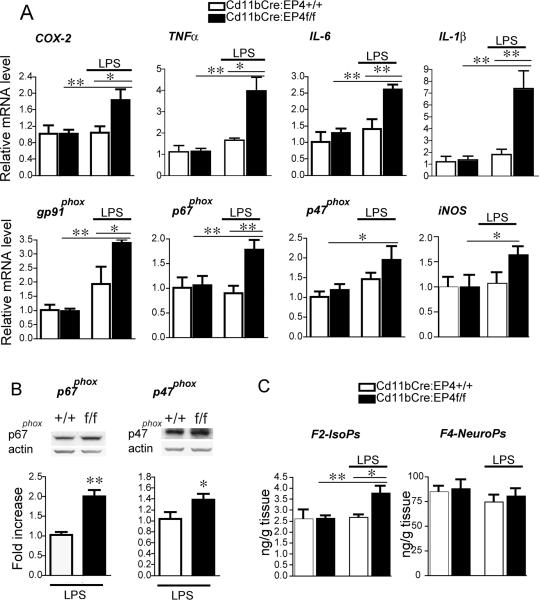
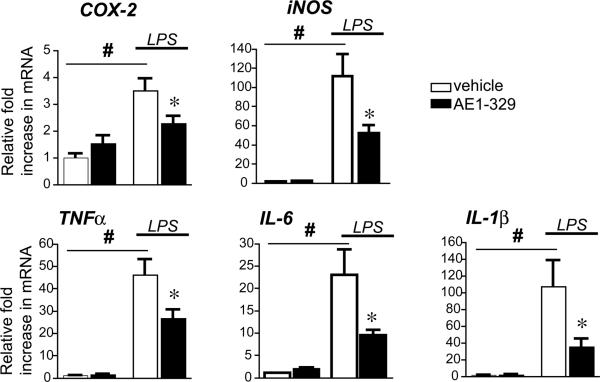
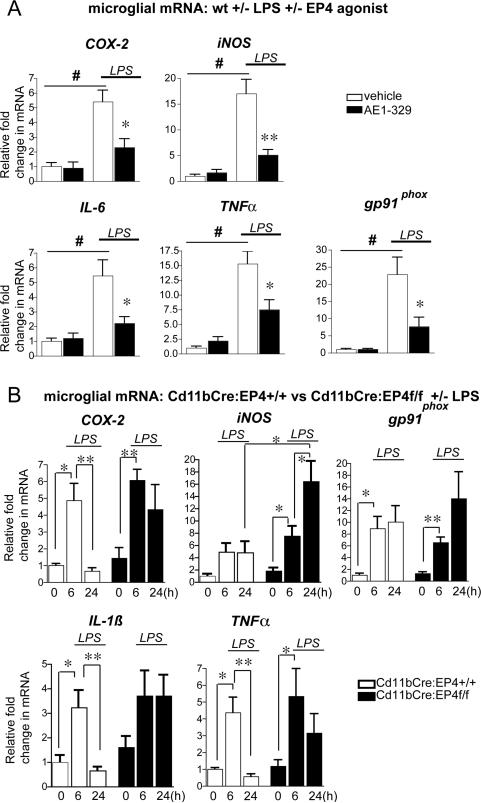
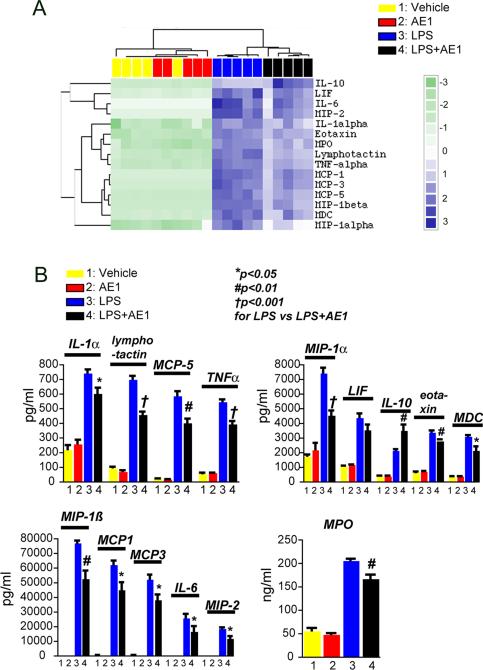
Peripheral inflammation leads to immune responses in brain characterized by microglial activation, elaboration of proinflammatory cytokines and reactive oxygen species, and secondary neuronal injury. The inducible cyclooxygenase (COX), COX-2, mediates a significant component of this response in brain via downstream proinflammatory PG signaling. In this study, we investigated the function of the PGE2 E-prostanoid (EP) 4 receptor in the CNS innate immune response to the bacterial endotoxin LPS. We report that PGE2 EP4 signaling mediates an anti-inflammatory effect in brain by blocking LPS-induced proinflammatory gene expression in mice. This was associated in cultured murine microglial cells with decreased Akt and I-kappaB kinase phosphorylation and decreased nuclear translocation of p65 and p50 NF-kappaB subunits. In vivo, conditional deletion of EP4 in macrophages and microglia increased lipid peroxidation and proinflammatory gene expression in brain and in isolated adult microglia following peripheral LPS administration. Conversely, EP4 selective agonist decreased LPS-induced proinflammatory gene expression in hippocampus and in isolated adult microglia. In plasma, EP4 agonist significantly reduced levels of proinflammatory cytokines and chemokines, indicating that peripheral EP4 activation protects the brain from systemic inflammation. The innate immune response is an important component of disease progression in a number of neurodegenerative disorders, such as Alzheimer's disease, Parkinson's disease, and amyotrophic lateral sclerosis. In addition, recent studies demonstrated adverse vascular effects with chronic administration of COX-2 inhibitors, indicating that specific PG signaling pathways may be protective in vascular function. This study supports an analogous and beneficial effect of PGE2 EP4 receptor signaling in suppressing brain inflammation.
Figures
Similar articles
-
Differential expression of E-type prostanoid receptors 2 and 4 in microglia stimulated with lipopolysaccharide.J Neuroinflammation. 2017 Jan 5;14(1):3. doi: 10.1186/s12974-016-0780-7. J Neuroinflammation. 2017. PMID: 28086956 Free PMC article.
-
Suppression of Alzheimer-associated inflammation by microglial prostaglandin-E2 EP4 receptor signaling.J Neurosci. 2014 Apr 23;34(17):5882-94. doi: 10.1523/JNEUROSCI.0410-14.2014. J Neurosci. 2014. PMID: 24760848 Free PMC article.
-
LXW7 attenuates inflammation via suppressing Akt/nuclear factor kappa B and mitogen-activated protein kinases signaling pathways in lipopolysaccharide-stimulated BV2 microglial cells.Int Immunopharmacol. 2019 Dec;77:105963. doi: 10.1016/j.intimp.2019.105963. Epub 2019 Nov 13. Int Immunopharmacol. 2019. PMID: 31732449
-
Prostaglandin E2-induced inflammation: Relevance of prostaglandin E receptors.Biochim Biophys Acta. 2015 Apr;1851(4):414-21. doi: 10.1016/j.bbalip.2014.07.008. Epub 2014 Jul 17. Biochim Biophys Acta. 2015. PMID: 25038274 Review.
-
Anti-inflammation therapy by activation of prostaglandin EP4 receptor in cardiovascular and other inflammatory diseases.J Cardiovasc Pharmacol. 2012 Feb;59(2):116-23. doi: 10.1097/FJC.0b013e3182244a12. J Cardiovasc Pharmacol. 2012. PMID: 21697732 Free PMC article. Review.
Cited by
-
Role of prostaglandins in neuroinflammatory and neurodegenerative diseases.Mediators Inflamm. 2012;2012:946813. doi: 10.1155/2012/946813. Epub 2012 Jun 18. Mediators Inflamm. 2012. PMID: 22778499 Free PMC article. Review.
-
Therapeutic implications of the prostaglandin pathway in Alzheimer's disease.Biochem Pharmacol. 2014 Apr 15;88(4):565-72. doi: 10.1016/j.bcp.2013.12.014. Epub 2014 Jan 13. Biochem Pharmacol. 2014. PMID: 24434190 Free PMC article. Review.
-
Astrocytic TGF-β signaling limits inflammation and reduces neuronal damage during central nervous system Toxoplasma infection.J Immunol. 2014 Jul 1;193(1):139-49. doi: 10.4049/jimmunol.1303284. Epub 2014 May 23. J Immunol. 2014. PMID: 24860191 Free PMC article.
-
Prostaglandins in the Inflamed Central Nervous System: Potential Therapeutic Targets.Curr Drug Targets. 2024;25(13):885-908. doi: 10.2174/0113894501323980240815113851. Curr Drug Targets. 2024. PMID: 39177131 Free PMC article. Review.
-
Receptors on Microglia.Adv Neurobiol. 2024;37:83-121. doi: 10.1007/978-3-031-55529-9_6. Adv Neurobiol. 2024. PMID: 39207688 Review.
References
-
- Nguyen MD, Julien JP, Rivest S. Innate immunity: the missing link in neuroprotection and neurodegeneration? Nature reviews. 2002;3:216–227. - PubMed
-
- Fassbender K, Walter S, Kuhl S, Landmann R, Ishii K, Bertsch T, Stalder AK, Muehlhauser F, Liu Y, Ulmer AJ, Rivest S, Lentschat A, Gulbins E, Jucker M, Staufenbiel M, Brechtel K, Walter J, Multhaup G, Penke B, Adachi Y, Hartmann T, Beyreuther K. The LPS receptor (CD14) links innate immunity with Alzheimer's disease. Faseb J. 2004;18:203–205. - PubMed
-
- Walter S, Letiembre M, Liu Y, Heine H, Penke B, Hao W, Bode B, Manietta N, Walter J, Schulz-Schuffer W, Fassbender K. Role of the toll-like receptor 4 in neuroinflammation in Alzheimer's disease. Cell Physiol Biochem. 2007;20:947–956. - PubMed
-
- Moisse K, Strong MJ. Innate immunity in amyotrophic lateral sclerosis. Biochim Biophys Acta. 2006;1762:1083–1093. - PubMed
-
- Letiembre M, Liu Y, Walter S, Hao W, Pfander T, Wrede A, Schulz-Schaeffer W, Fassbender K. Screening of innate immune receptors in neurodegenerative diseases: A similar pattern. Neurobiol Aging. 2007 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
