

The alternative end-joining pathway for repair of DNA double-strand breaks requires PARP1 but is not dependent upon microhomologies
- PMID: 20483915
- PMCID: PMC2952854
- DOI: 10.1093/nar/gkq387
The alternative end-joining pathway for repair of DNA double-strand breaks requires PARP1 but is not dependent upon microhomologies
Abstract
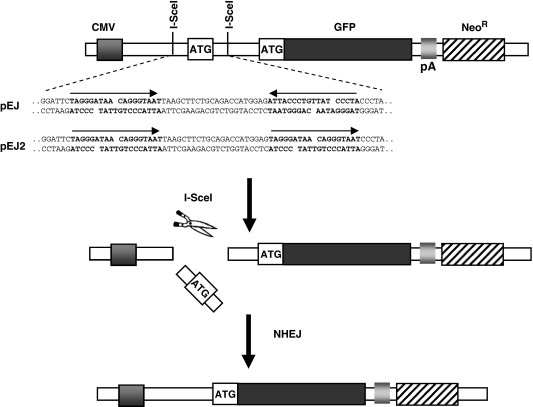
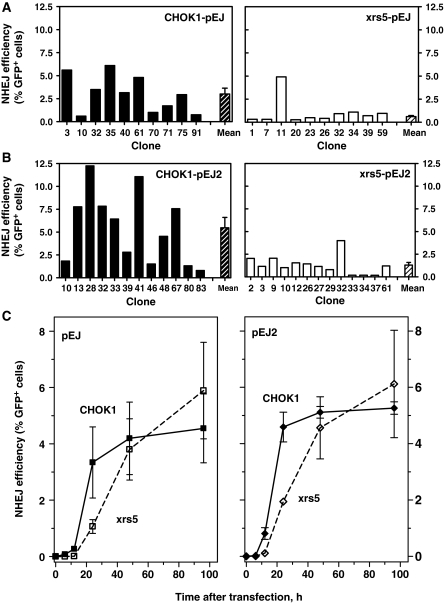
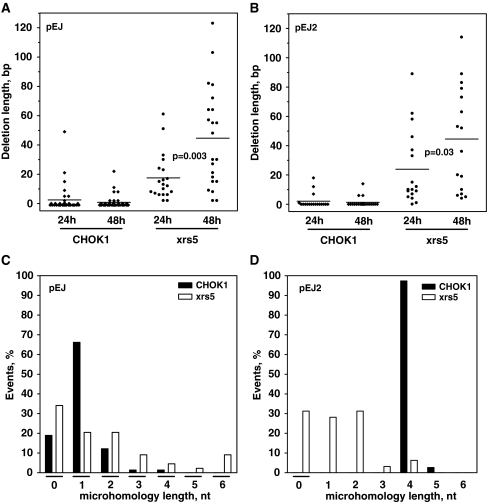
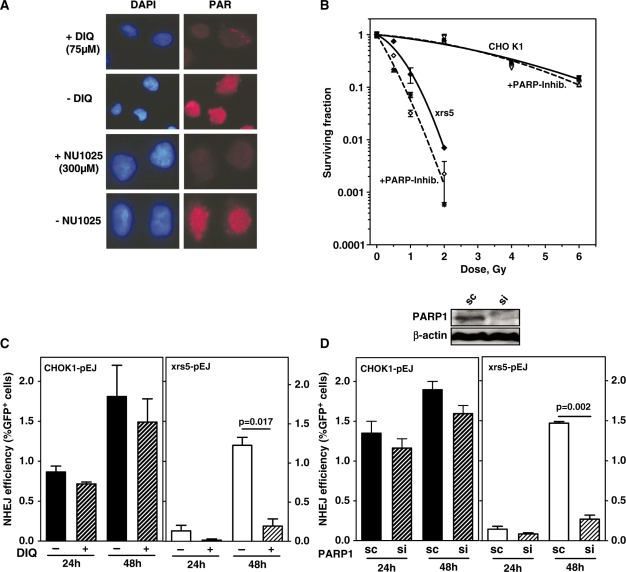
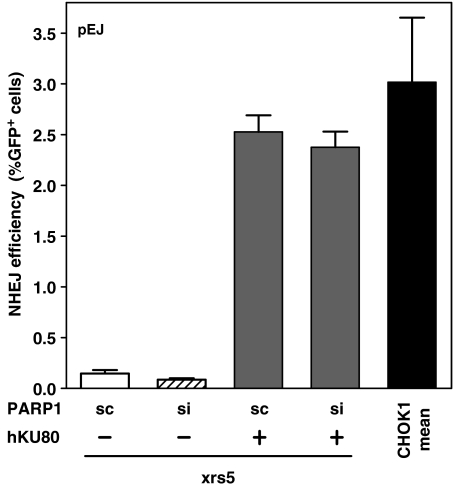
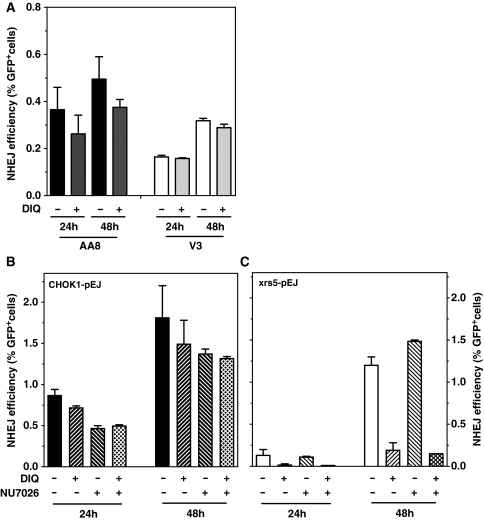
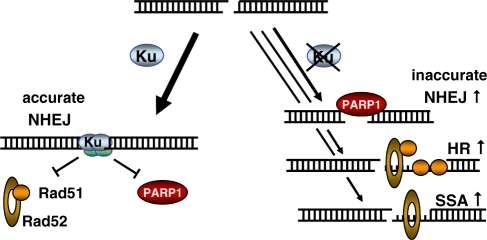
Non-homologous end-joining (NHEJ), the major repair pathway for DNA double-strand breaks (DSB) in mammalian cells, employs a repertoire of core proteins, the recruitment of which to DSB-ends is Ku-dependent. Lack of either of the core components invariably leads to a repair deficiency. There has been evidence that an alternative end-joining operates in the absence of the core components. We used chromosomal reporter substrates to specifically monitor NHEJ of single I-SceI-induced-DSB for detailed comparison of classical and alternative end-joining. We show that rapid repair of both compatible and non-compatible ends require Ku-protein. In the absence of Ku, cells use a slow but efficient repair mode which experiences increasing sequence-loss with time after DSB induction. Chemical inhibition and PARP1-depletion demonstrated that the alternative end-joining in vivo is completely dependent upon functional PARP1. Furthermore, we show that the requirement for PARP1 depends on the absence of Ku but not on DNA-dependent protein kinase (DNA-PKcs). Extensive sequencing of repair junctions revealed that the alternative rejoining does not require long microhomologies. Together, we show that mammalian cells need Ku for rapid and conservative NHEJ. PARP1-dependent alternative route may partially rescue the deficient repair phenotype presumably at the expense of an enhanced mutation rate.
Figures
Similar articles
-
The absence of Ku but not defects in classical non-homologous end-joining is required to trigger PARP1-dependent end-joining.DNA Repair (Amst). 2013 Dec;12(12):1134-42. doi: 10.1016/j.dnarep.2013.10.005. Epub 2013 Nov 7. DNA Repair (Amst). 2013. PMID: 24210699
-
PARP-1 and Ku compete for repair of DNA double strand breaks by distinct NHEJ pathways.Nucleic Acids Res. 2006;34(21):6170-82. doi: 10.1093/nar/gkl840. Epub 2006 Nov 6. Nucleic Acids Res. 2006. PMID: 17088286 Free PMC article.
-
CRISPR/Cas9-Induced Double-Strand Break Repair in Arabidopsis Nonhomologous End-Joining Mutants.G3 (Bethesda). 2017 Jan 5;7(1):193-202. doi: 10.1534/g3.116.035204. G3 (Bethesda). 2017. PMID: 27866150 Free PMC article.
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
-
[Double strand break repair, one mechanism can hide another: alternative non-homologous end joining].Cancer Radiother. 2012 Feb;16(1):1-10. doi: 10.1016/j.canrad.2011.05.004. Epub 2011 Jul 6. Cancer Radiother. 2012. PMID: 21737335 Review. French.
Cited by
-
Establishment of a Transformation Coupled in vitro End Joining Assay to Estimate Radiosensitivity in Tumor Cells.Front Oncol. 2020 Aug 20;10:1480. doi: 10.3389/fonc.2020.01480. eCollection 2020. Front Oncol. 2020. PMID: 32974177 Free PMC article.
-
Chemical and biological approaches to improve the efficiency of homologous recombination in human cells mediated by artificial restriction DNA cutter.Nucleic Acids Res. 2012 Jun;40(11):e81. doi: 10.1093/nar/gks185. Epub 2012 Feb 23. Nucleic Acids Res. 2012. PMID: 22362741 Free PMC article.
-
The expanding universe of PARP1-mediated molecular and therapeutic mechanisms.Mol Cell. 2022 Jun 16;82(12):2315-2334. doi: 10.1016/j.molcel.2022.02.021. Epub 2022 Mar 9. Mol Cell. 2022. PMID: 35271815 Free PMC article. Review.
-
ATM-dependent phosphorylation of CHD7 regulates morphogenesis-coupled DSB stress response in fetal radiation exposure.Mol Biol Cell. 2023 May 1;34(5):ar39. doi: 10.1091/mbc.E22-10-0450. Epub 2023 Mar 8. Mol Biol Cell. 2023. PMID: 36884301 Free PMC article.
-
PARP Inhibitors: A New Horizon for Patients with Prostate Cancer.Biomedicines. 2022 Jun 15;10(6):1416. doi: 10.3390/biomedicines10061416. Biomedicines. 2022. PMID: 35740437 Free PMC article. Review.
References
-
- Lieber MR. The mechanism of human nonhomologous DNA end joining. J. Biol. Chem. 2008;283:1–5. - PubMed
-
- Ma Y, Schwarz K, Lieber MR. The Artemis: DNA-PKcs endonuclease cleaves DNA loops, flaps, and gaps. DNA Repair. 2005;4:845–851. - PubMed
-
- Audebert M, Salles B, Weinfeld M, Calsou P. Involvement of polynucleotide kinase in a poly(ADP-ribose) polymerase-1-dependent DNA double-strand breaks rejoining pathway. J. Mol. Biol. 2006;356:257–265. - PubMed
-
- Jeggo PA, Kemp LM. X-ray-sensitive mutants of Chinese hamster ovary cell line. Isolation and cross-sensitivity to other DNA-damaging agents. Mutat. Res. 1983;112:313–327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
