

Structural basis on the dityrosyl-diiron radical cluster and the functional differences of human ribonucleotide reductase small subunits hp53R2 and hRRM2
- PMID: 20484015
- PMCID: PMC3050530
- DOI: 10.1158/1535-7163.MCT-10-0023
Structural basis on the dityrosyl-diiron radical cluster and the functional differences of human ribonucleotide reductase small subunits hp53R2 and hRRM2
Abstract
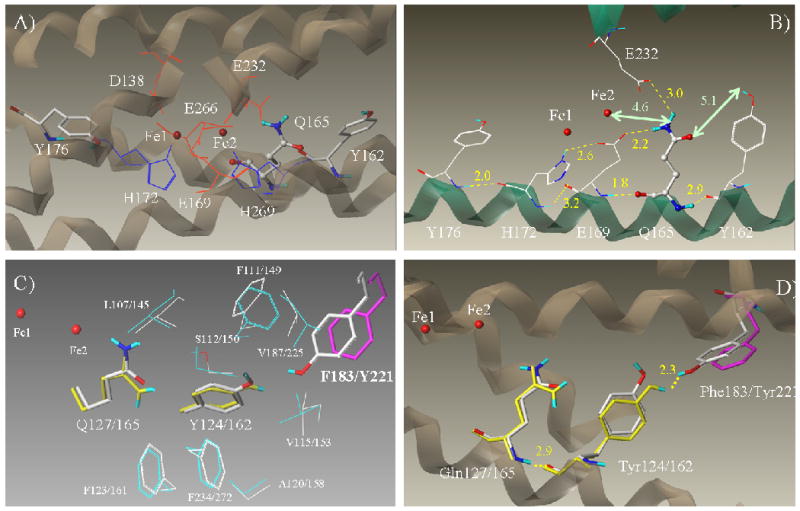
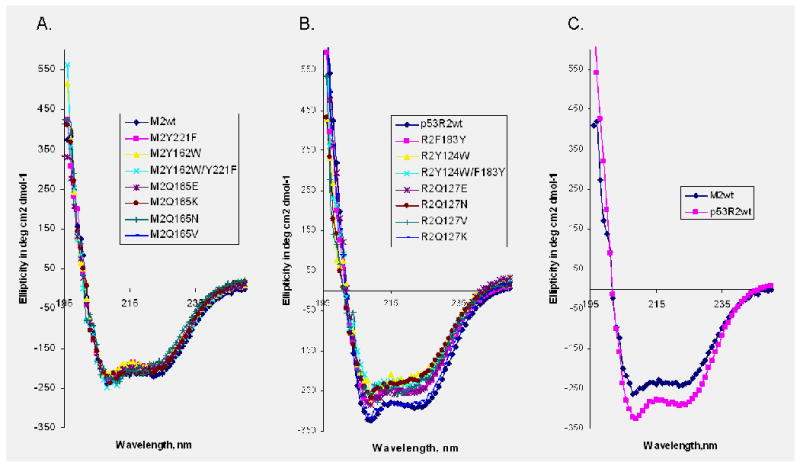
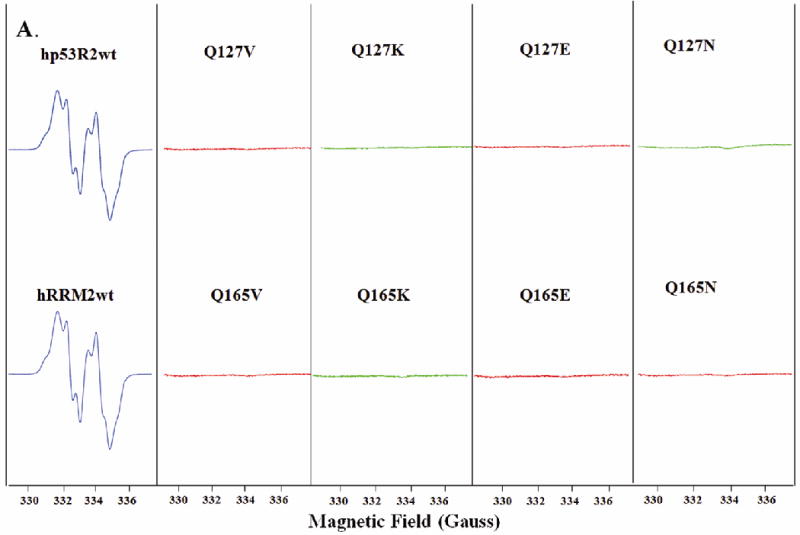
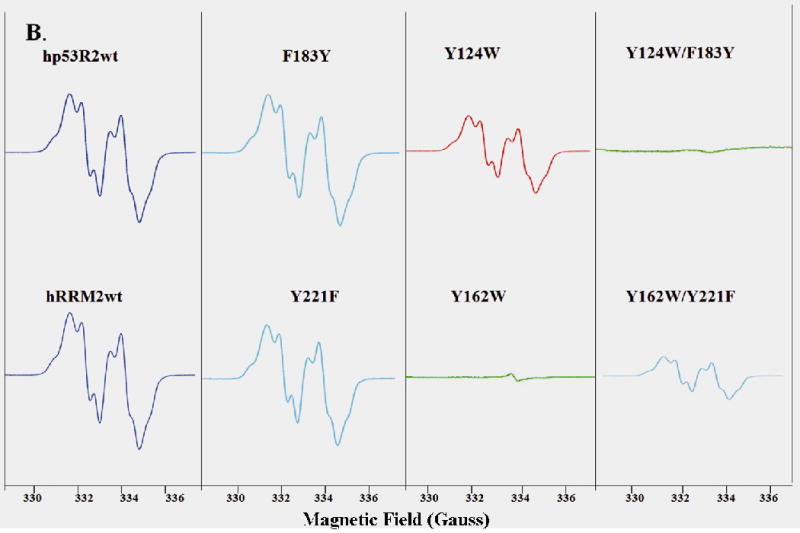
Ribonucleotide reductase (RNR) is an enzyme for the de novo conversion of ribonucleotides to deoxyribonucleotides. The two human RNR small subunits hRRM2 and hp53R2 share 83% sequence homology but show distinct expression patterns and function. Structural analyses of the oxidized form of hRRM2 and hp53R2 indicate that both proteins contain a conserved Gln127-hp53R2/Gln165-hRRM2 close to the dinuclear iron center and the essential tyrosine residue Tyr124-hp53R2/Tyr162-hRRM2 forms hydrogen bonds with the tyrosine and iron ligands, implying a critical role for the glutamine residue in assembling the dityrosyl-diiron radical cofactor. The present work also showed that Tyr221 in hRRM2, which is replaced by Phe183 in hp53R2, forms a hydrogen bond with Tyr162 to extend the hydrogen bond network from Gln165-hRRM2. Mutagenesis and spectroscopic experiments suggested that the tyrosine-to-phenylalanine switch at Phe183-hp53R2/Tyr221-hRRM2 could lead to differences in radical generation or enzymatic activity for hp53R2 and hRRM2. This study correlates the distinct catalytic mechanisms of the small subunits hp53R2 and hRRM2 with a hydrogen-bonding network and provides novel directions for designing and developing subunit-specific therapeutic agents for human RNR enzymes.
Conflict of interest statement
No potential conflicts of interest were disclosed
Figures
References
-
- Eklund H, Uhlin U, Färnegårdh M, Logan DT, Nordlund P. Structure and function of the radical enzyme ribonucleotide reductase. Prog Biophys Mol Biol. 2001;77:177–268. - PubMed
-
- Kolberg M, Strand KR, Graff R, Andersson KK. Structure, function, and mechanism of ribonucleotide reductases. Biochim Biophys Acta. 2004;1699:1–34. - PubMed
-
- Sjöberg BM. Ribonucleotide reductases - A group of enzymes with different metallosites and a similar reaction mechanism. Struct Bonding. 1997;88:139.
-
- Stubbe J, Nocera DG, Yee CS, Chang MCY. Radical initiation in the class I ribonucleotide reductase: Long-range proton-coupled electron transfer? Chem Rev. 2003;103:2167–2201. - PubMed
-
- Gon S, Beckwith J. Ribonucleotide reductases: influence of environment on synthesis and activity. Antioxid Redox Signal. 2006;8:773–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
