

TREM2- and DAP12-dependent activation of PI3K requires DAP10 and is inhibited by SHIP1
- PMID: 20484116
- PMCID: PMC2900152
- DOI: 10.1126/scisignal.2000500
TREM2- and DAP12-dependent activation of PI3K requires DAP10 and is inhibited by SHIP1
Abstract
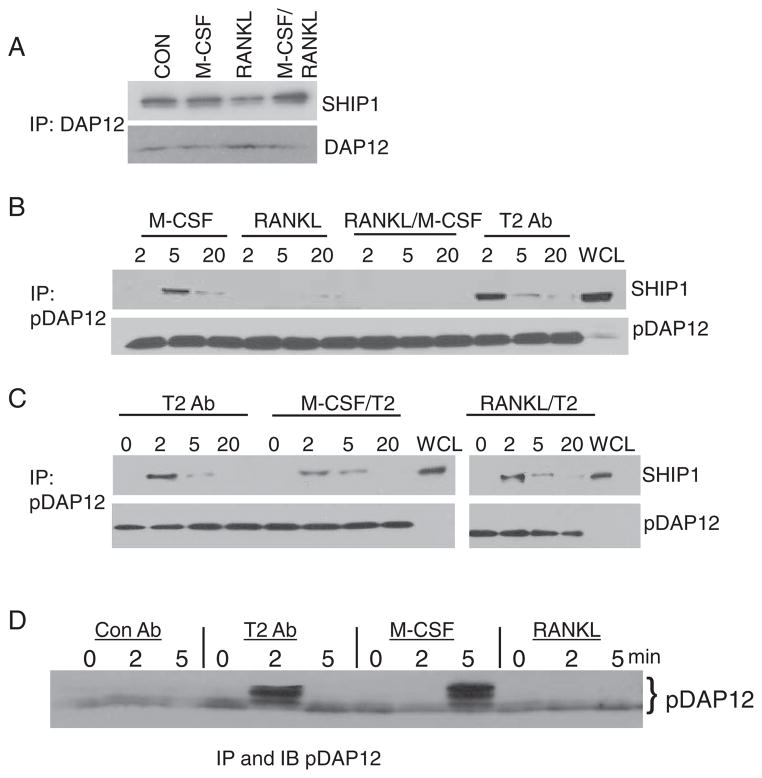
The activation and fusion of macrophages and of osteoclasts require the adaptor molecule DNAX-activating protein of 12 kD (DAP12), which contains immunoreceptor tyrosine-based activation motifs (ITAMs). TREM2 (triggering receptor expressed on myeloid cells-2) is the main DAP12-associated receptor in osteoclasts and, similar to DAP12 deficiency, loss of TREM2 in humans leads to Nasu-Hakola disease, which is characterized by bone cysts and dementia. Furthermore, in vitro experiments have shown that deficiency in DAP12 or TREM2 leads to impaired osteoclast development and the formation of mononuclear osteoclasts. Here, we demonstrate that the ligation of TREM2 activated phosphatidylinositol 3-kinase (PI3K), extracellular signal-regulated kinase 1 (ERK1) and ERK2, and the guanine nucleotide exchange factor Vav3; induced the mobilization of intracellular calcium (Ca(2+)) and the reorganization of actin; and prevented apoptosis. The signaling adaptor molecule DAP10 played a key role in the TREM2- and DAP12-dependent recruitment of PI3K to the signaling complex. Src homology 2 (SH2) domain-containing inositol phosphatase-1 (SHIP1) inhibited TREM2- and DAP12-induced signaling by binding to DAP12 in an SH2 domain-dependent manner and preventing the recruitment of PI3K to DAP12. These results demonstrate a previously uncharacterized interaction of SHIP1 with DAP12 that functionally limits TREM2- and DAP12-dependent signaling and identify a mechanism through which SHIP1 regulates key ITAM-containing receptors by directly blocking the binding and activation of PI3K.
Conflict of interest statement
Figures
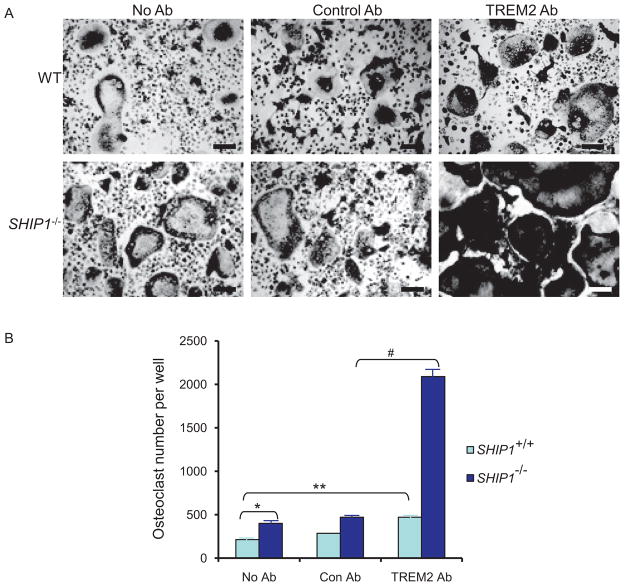
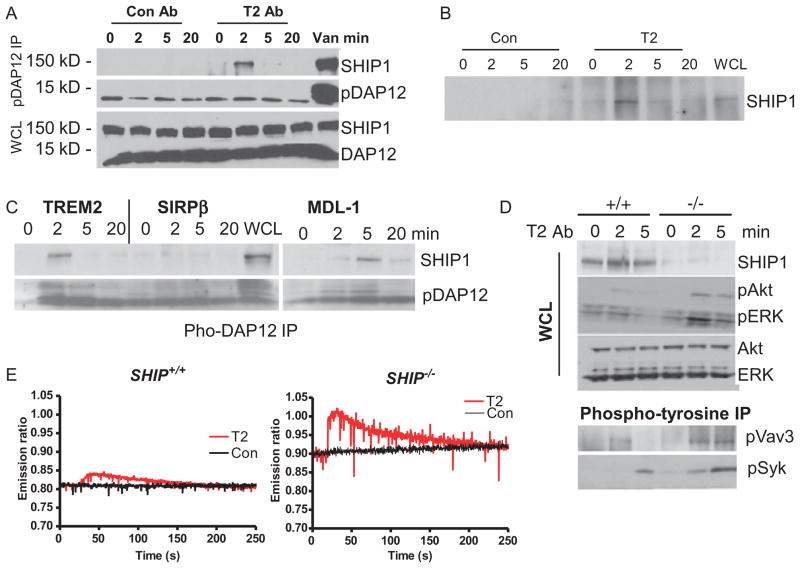

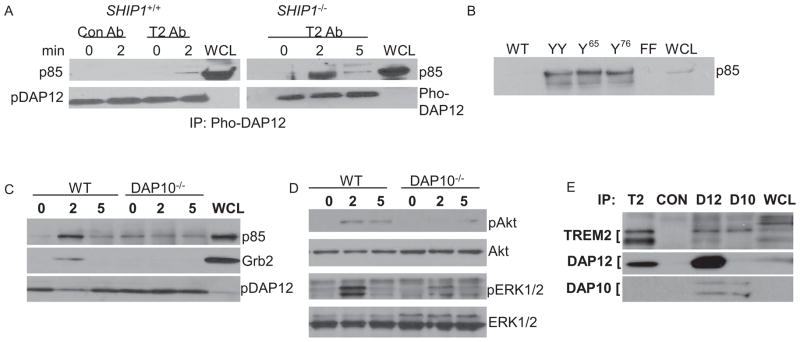
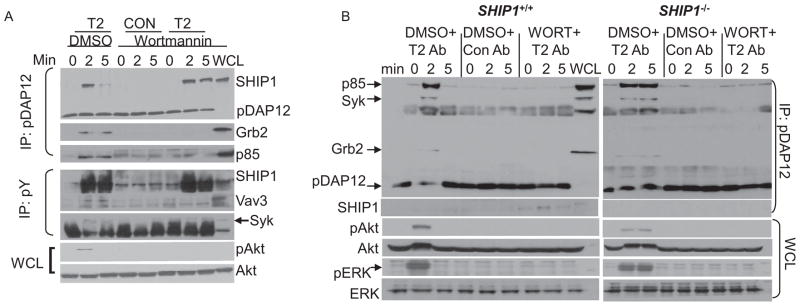



References
-
- Mócsai A, Humphrey MB, Van Ziffle JA, Hu Y, Burghardt A, Spusta SC, Majumdar S, Lanier LL, Lowell CA, Nakamura MC. The immunomodulatory adapter proteins DAP12 and Fc receptor γ-chain (FcRγ) regulate development of functional osteoclasts through the Syk tyrosine kinase. Proc Natl Acad Sci USA. 2004;101:6158–6163. - PMC - PubMed
-
- Humphrey MB, Ogasawara K, Yao W, Spusta SC, Daws MR, Lane NE, Lanier LL, Nakamura MC. The signaling adapter protein DAP12 regulates multinucleation during osteoclast development. J Bone Miner Res. 2004;19:224–234. - PubMed
-
- Koga T, Inui M, Inoue K, Kim S, Suematsu A, Kobayashi E, Iwata T, Ohnishi H, Matozaki T, Kodama T, Taniguchi T, Takayanagi H, Takai T. Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature. 2004;428:758–763. - PubMed
-
- Nataf S, Anginot A, Vuaillat C, Malaval L, Fodil N, Chereul E, Langlois JB, Dumontel C, Cavillon G, Confavreux C, Mazzorana M, Vico L, Belin MF, Vivier E, Tomasello E, Jurdic P. Brain and bone damage in KARAP/DAP12 loss-of-function mice correlate with alterations in microglia and osteoclast lineages. Am J Pathol. 2005;166:275–286. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
