

The ubiquitin-proteasome system plays an important role during various stages of the coronavirus infection cycle
- PMID: 20484504
- PMCID: PMC2897594
- DOI: 10.1128/JVI.00485-10
The ubiquitin-proteasome system plays an important role during various stages of the coronavirus infection cycle
Abstract
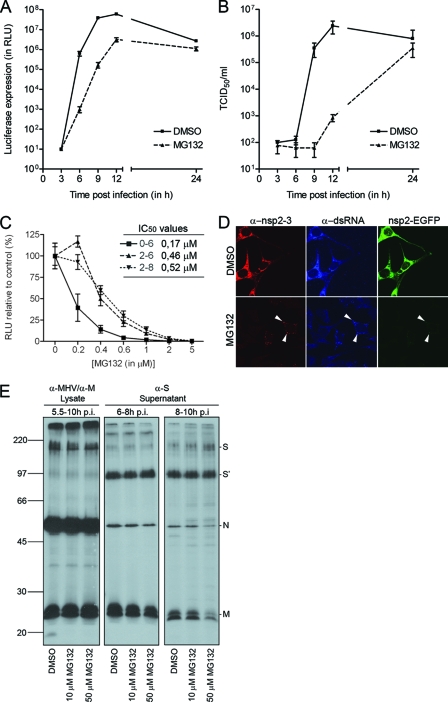
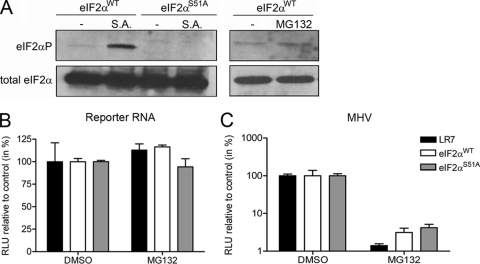
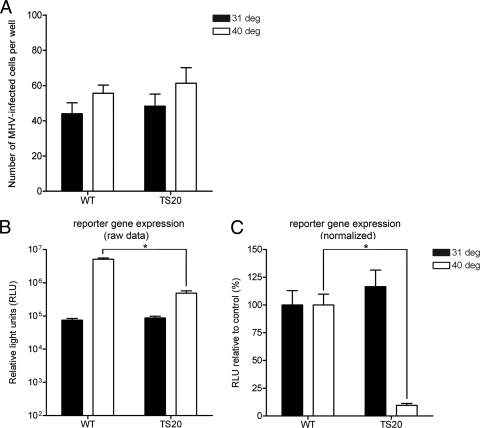
The ubiquitin-proteasome system (UPS) is a key player in regulating the intracellular sorting and degradation of proteins. In this study we investigated the role of the UPS in different steps of the coronavirus (CoV) infection cycle. Inhibition of the proteasome by different chemical compounds (i.e., MG132, epoxomicin, and Velcade) appeared to not only impair entry but also RNA synthesis and subsequent protein expression of different CoVs (i.e., mouse hepatitis virus [MHV], feline infectious peritonitis virus, and severe acute respiratory syndrome CoV). MHV assembly and release were, however, not appreciably affected by these compounds. The inhibitory effect on CoV protein expression did not appear to result from a general inhibition of translation due to induction of a cellular stress response by the inhibitors. Stress-induced phosphorylation of eukaryotic translation initiation factor 2alpha (eIF2alpha) generally results in impaired initiation of protein synthesis, but the sensitivity of MHV infection to proteasome inhibitors was unchanged in cells lacking a phosphorylatable eIF2alpha. MHV infection was affected not only by inhibition of the proteasome but also by interfering with protein ubiquitination. Viral protein expression was reduced in cells expressing a temperature-sensitive ubiquitin-activating enzyme E1 at the restrictive temperature, as well as in cells in which ubiquitin was depleted by using small interfering RNAs. Under these conditions, the susceptibility of the cells to virus infection was, however, not affected, excluding an important role of ubiquitination in virus entry. Our observations reveal an important role of the UPS in multiple steps of the CoV infection cycle and identify the UPS as a potential drug target to modulate the impact of CoV infection.
Figures



References
-
- Anand, K., J. Ziebuhr, P. Wadhwani, J. R. Mesters, and R. Hilgenfeld. 2003. Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs. Science 300:1763-1767. - PubMed
-
- Berlanga, J. J., J. Santoyo, and C. De Haro. 1999. Characterization of a mammalian homolog of the GCN2 eukaryotic initiation factor 2α kinase. Eur. J. Biochem. 265:754-762. - PubMed
-
- Bieniasz, P. D. 2006. Late budding domains and host proteins in enveloped virus release. Virology 344:55-63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
