

Temporal patterns of inputs to cerebellum necessary and sufficient for trace eyelid conditioning
- PMID: 20484534
- PMCID: PMC2934927
- DOI: 10.1152/jn.00169.2010
Temporal patterns of inputs to cerebellum necessary and sufficient for trace eyelid conditioning
Abstract
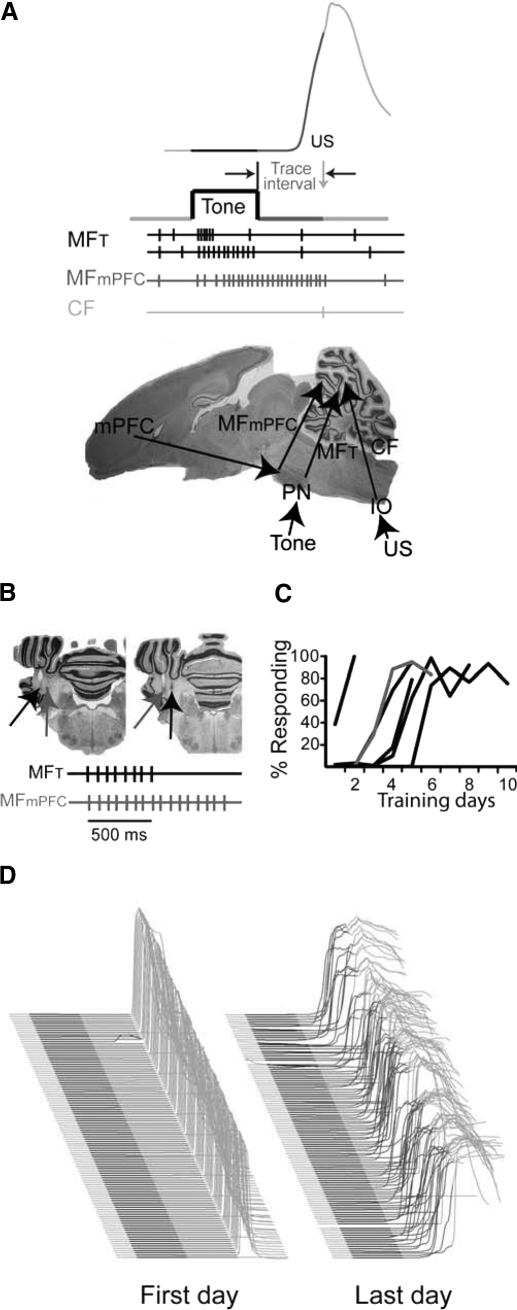
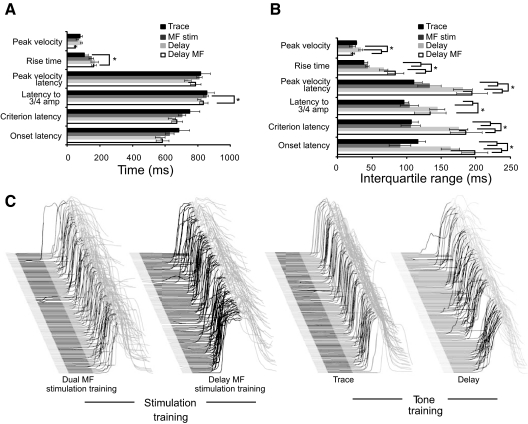
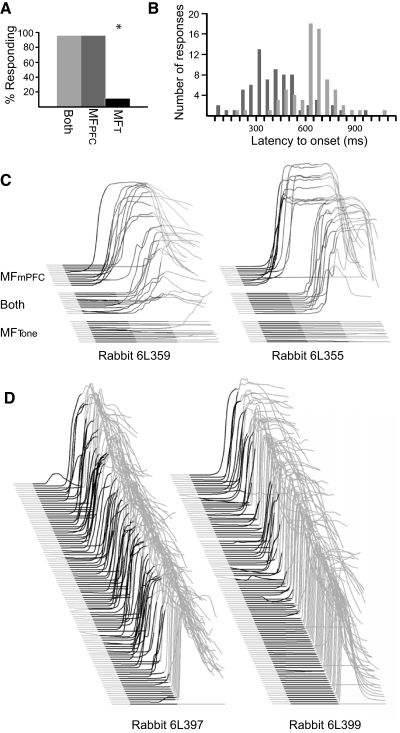
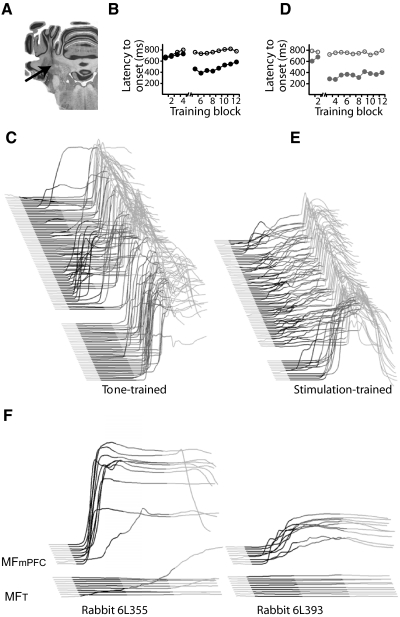
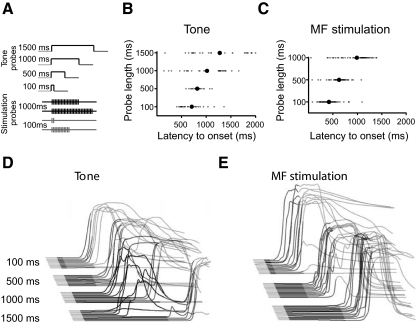
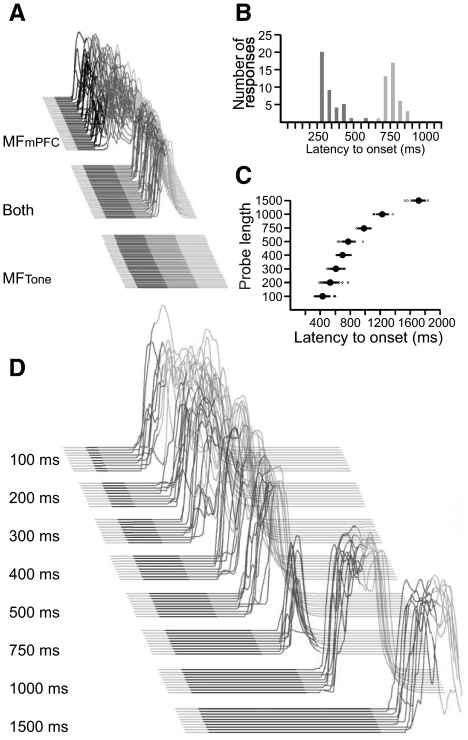
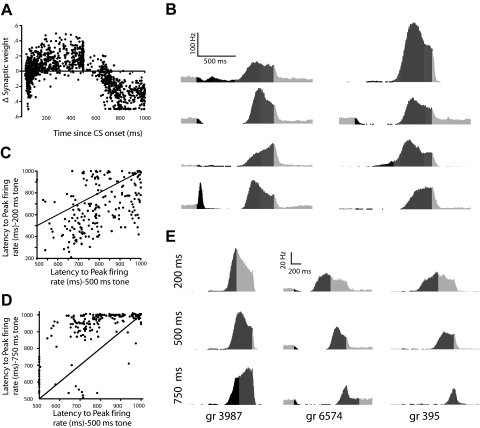
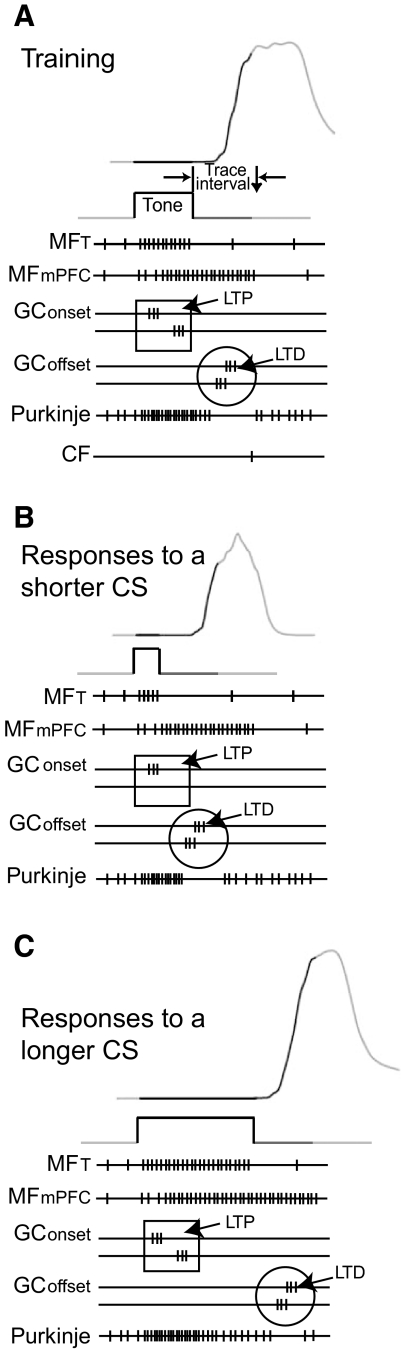
Trace eyelid conditioning is a form of associative learning that requires several forebrain structures and cerebellum. Previous work suggests that at least two conditioned stimulus (CS)-driven signals are available to the cerebellum via mossy fiber inputs during trace conditioning: one driven by and terminating with the tone and a second driven by medial prefrontal cortex (mPFC) that persists through the stimulus-free trace interval to overlap in time with the unconditioned stimulus (US). We used electric stimulation of mossy fibers to determine whether this pattern of dual inputs is necessary and sufficient for cerebellar learning to express normal trace eyelid responses. We find that presenting the cerebellum with one input that mimics persistent activity observed in mPFC and the lateral pontine nuclei during trace eyelid conditioning and another that mimics tone-elicited mossy fiber activity is sufficient to produce responses whose properties quantitatively match trace eyelid responses using a tone. Probe trials with each input delivered separately provide evidence that the cerebellum learns to respond to the mPFC-like input (that overlaps with the US) and learns to suppress responding to the tone-like input (that does not). This contributes to precisely timed responses and the well-documented influence of tone offset on the timing of trace responses. Computer simulations suggest that the underlying cerebellar mechanisms involve activation of different subsets of granule cells during the tone and during the stimulus-free trace interval. These results indicate that tone-driven and mPFC-like inputs are necessary and sufficient for the cerebellum to learn well-timed trace conditioned responses.
Figures
Similar articles
-
Persistent activity in prefrontal cortex during trace eyelid conditioning: dissociating responses that reflect cerebellar output from those that do not.J Neurosci. 2013 Sep 18;33(38):15272-84. doi: 10.1523/JNEUROSCI.1238-13.2013. J Neurosci. 2013. PMID: 24048856 Free PMC article.
-
Interactions between prefrontal cortex and cerebellum revealed by trace eyelid conditioning.Learn Mem. 2009 Jan 7;16(1):86-95. doi: 10.1101/lm.1178309. Print 2009 Jan. Learn Mem. 2009. PMID: 19144967 Free PMC article.
-
Persistent activity in a cortical-to-subcortical circuit: bridging the temporal gap in trace eyelid conditioning.J Neurophysiol. 2012 Jan;107(1):50-64. doi: 10.1152/jn.00689.2011. Epub 2011 Sep 28. J Neurophysiol. 2012. PMID: 21957220 Free PMC article.
-
Temporal discrimination in the cerebellar cortex during conditioned eyelid responses.Exp Brain Res. 1998 Jul;121(2):115-24. doi: 10.1007/s002210050443. Exp Brain Res. 1998. PMID: 9696380 Review.
-
Where is the trace in trace conditioning?Trends Neurosci. 2008 Feb;31(2):105-12. doi: 10.1016/j.tins.2007.11.006. Epub 2008 Jan 15. Trends Neurosci. 2008. PMID: 18199490 Review.
Cited by
-
Optogenetic inhibition of ventral hippocampal neurons alleviates associative motor learning dysfunction in a rodent model of schizophrenia.PLoS One. 2019 Dec 31;14(12):e0227200. doi: 10.1371/journal.pone.0227200. eCollection 2019. PLoS One. 2019. PMID: 31891640 Free PMC article.
-
Persistent activity in prefrontal cortex during trace eyelid conditioning: dissociating responses that reflect cerebellar output from those that do not.J Neurosci. 2013 Sep 18;33(38):15272-84. doi: 10.1523/JNEUROSCI.1238-13.2013. J Neurosci. 2013. PMID: 24048856 Free PMC article.
-
Differential effects of the cannabinoid agonist WIN55,212-2 on delay and trace eyeblink conditioning.Behav Neurosci. 2013 Oct;127(5):694-702. doi: 10.1037/a0034210. Behav Neurosci. 2013. PMID: 24128358 Free PMC article.
-
Reevaluating the role of the hippocampus in delay eyeblink conditioning.PLoS One. 2013 Aug 9;8(8):e71249. doi: 10.1371/journal.pone.0071249. eCollection 2013. PLoS One. 2013. PMID: 23951119 Free PMC article.
-
Awareness is essential for differential delay eyeblink conditioning with soft-tone but not loud-tone conditioned stimuli.Neurosci Bull. 2014 Jun;30(3):433-40. doi: 10.1007/s12264-013-1400-5. Epub 2014 Jan 29. Neurosci Bull. 2014. PMID: 24477990 Free PMC article.
References
-
- Aitkin LM, Boyd J. Acoustic input to the lateral pontine nuclei. Hear Res 1: 67–77, 1978 - PubMed
-
- Blum S, Hebert AE, Dash PK. A role for the prefrontal cortex in recall of recent and remote memories. Neuroreport 17: 341–344, 2006 - PubMed
-
- Boyd J, Aitkin LM. Responses of single units in the pontine nuclei of the cat to acoustic stimulation. Neurosci Lett 3: 259–263, 1976 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources