

Aurora-C kinase deficiency causes cytokinesis failure in meiosis I and production of large polyploid oocytes in mice
- PMID: 20484572
- PMCID: PMC2903667
- DOI: 10.1091/mbc.e10-02-0170
Aurora-C kinase deficiency causes cytokinesis failure in meiosis I and production of large polyploid oocytes in mice
Abstract
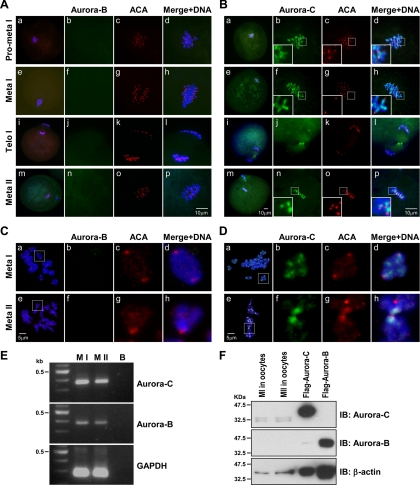
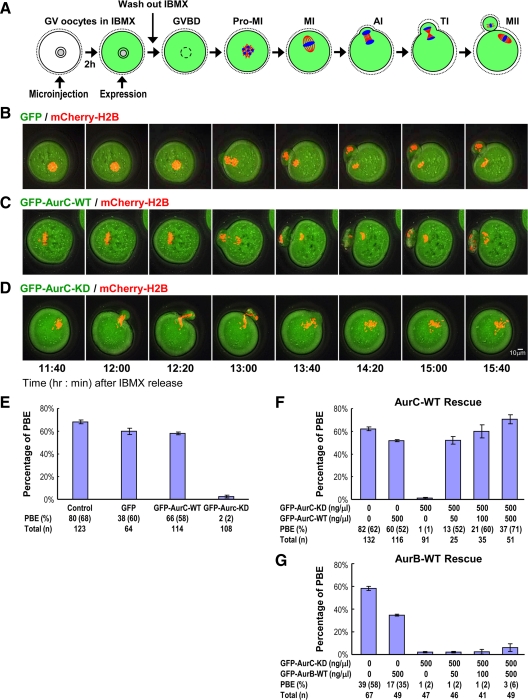
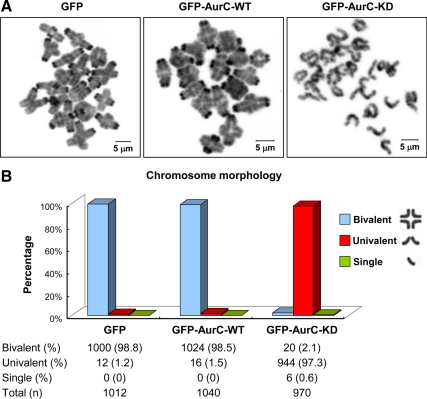
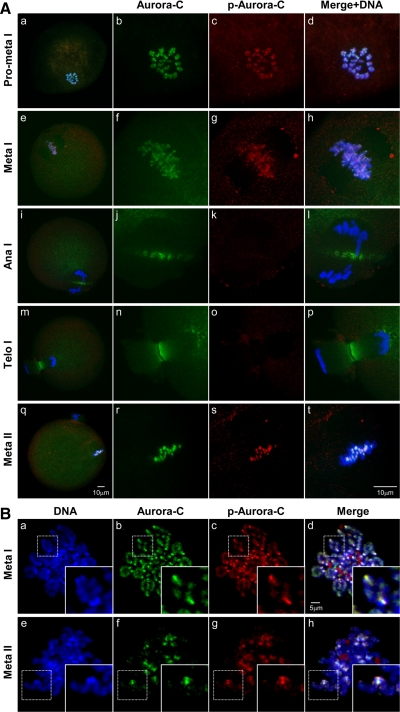
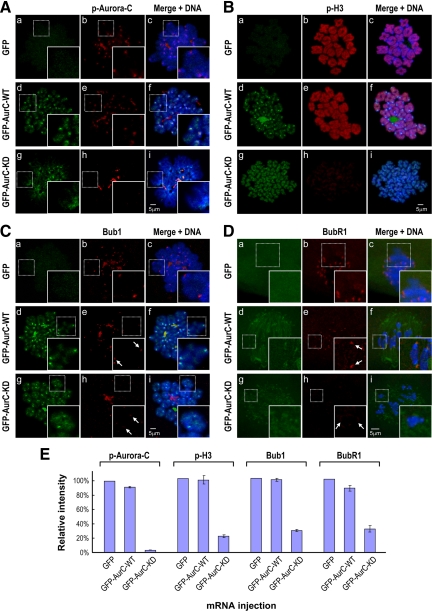
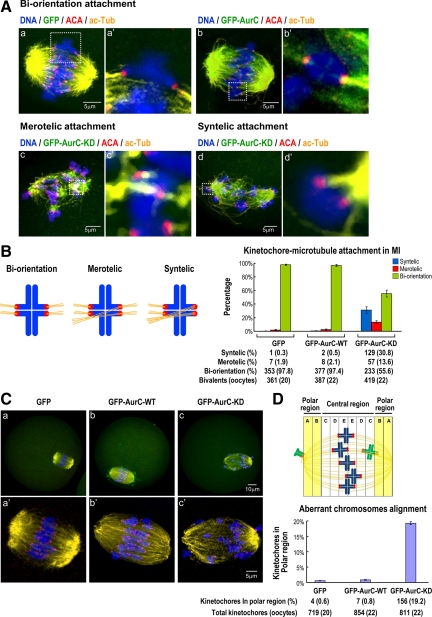
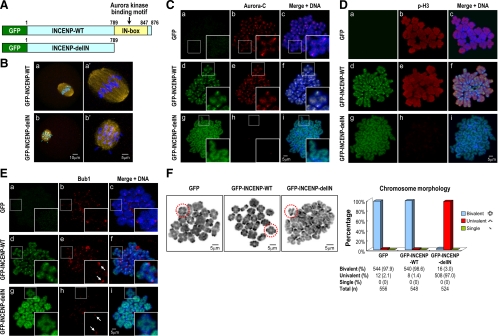
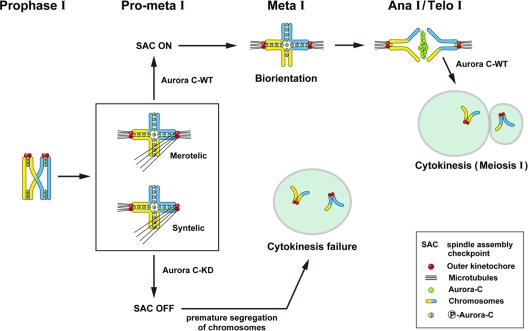
We previously isolated Aurora-C/Aie1 in a screen for kinases expressed in mouse sperm and eggs. Here, we show the localization of endogenous Aurora-C and examine its roles during female mouse meiosis. Aurora-C was detected at the centromeres and along the chromosome arms in prometaphase I-metaphase I and was concentrated at centromeres at metaphase II, in which Aurora-C also was phosphorylated at Thr171. During the anaphase I-telophase I transition, Aurora-C was dephosphorylated and relocalized to the midzone and midbody. Microinjection of the kinase-deficient Aurora-C (AurC-KD) mRNA into mouse oocytes significantly inhibited Aurora-C activity and caused multiple defects, including chromosome misalignment, abnormal kinetochore-microtubule attachment, premature chromosome segregation, and cytokinesis failure in meiosis I. Furthermore, AurC-KD reduced Aurora-C and histone H3 phosphorylation and inhibited kinetochore localization of Bub1 and BubR1. Similar effects also were observed in the oocytes injected with INCNEP-delIN mRNAs, in which the Aurora-C binding motif was removed. The most dramatic effect observed in AurC-KD-injected oocytes is cytokinesis failure in meiosis I, resulting in producing large polyploid oocytes, a pattern similar to Aurora-C deficiency human spermatozoa. Surprisingly, we detected no Aurora-B protein in mouse oocytes. We propose that Aurora-C, but not Aurora-B, plays essential roles in female mouse meiosis.
Figures
References
-
- Andrews P. D., Knatko E., Moore W. J., Swedlow J. R. Mitotic mechanics: the auroras come into view. Curr. Opin. Cell Biol. 2003;15:672–683. - PubMed
-
- Bernard M., Sanseau P., Henry C., Couturier A., Prigent C. Cloning of STK13, a third human protein kinase related to Drosophila aurora and budding yeast Ipl1 that maps on chromosome 19q13.3-ter. Genomics. 1998;53:406–409. - PubMed
-
- Carmena M., Earnshaw W. C. The cellular geography of aurora kinases. Nat. Rev. Mol. Cell Biol. 2003;4:842–854. - PubMed
-
- Chen H. L., Tang C. J., Chen C. Y., Tang T. K. Overexpression of an Aurora-C kinase-deficient mutant disrupts the Aurora-B/INCENP complex and induces polyploidy. J. Biomed. Sci. 2005;12:297–310. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
