

Connexin 43 mediates the tangential to radial migratory switch in ventrally derived cortical interneurons
- PMID: 20484649
- PMCID: PMC2883285
- DOI: 10.1523/JNEUROSCI.5728-09.2010
Connexin 43 mediates the tangential to radial migratory switch in ventrally derived cortical interneurons
Abstract
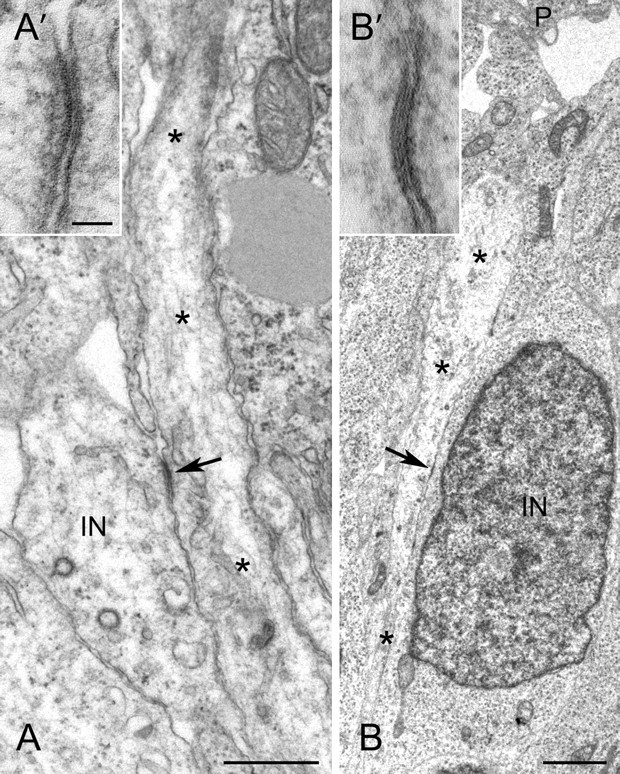
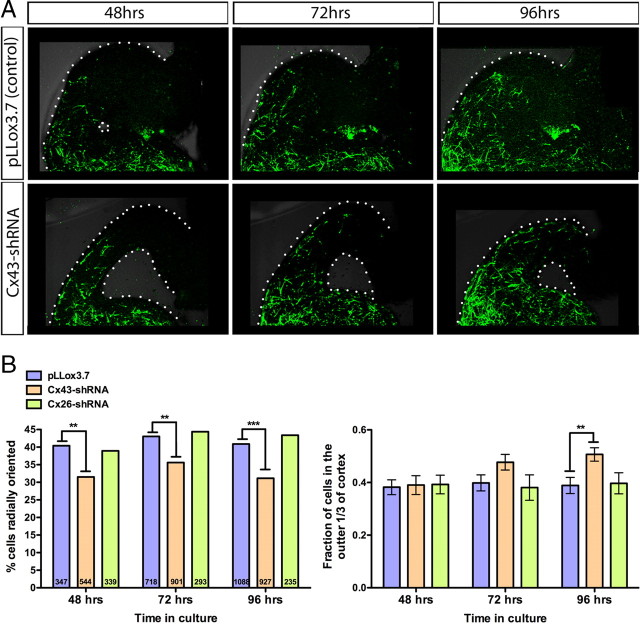
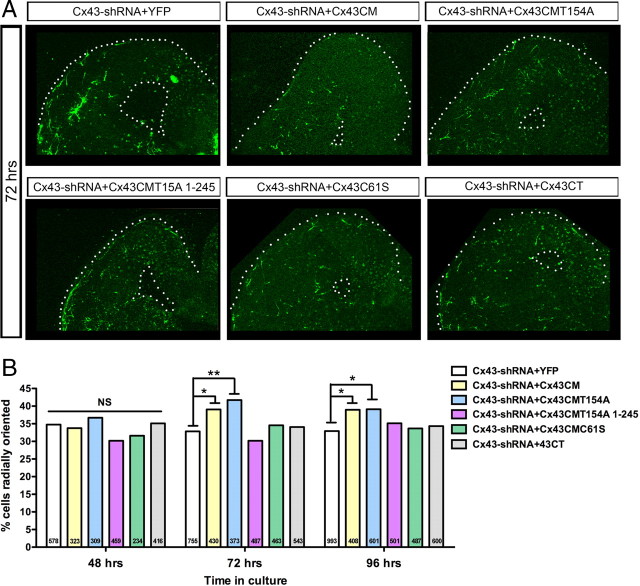
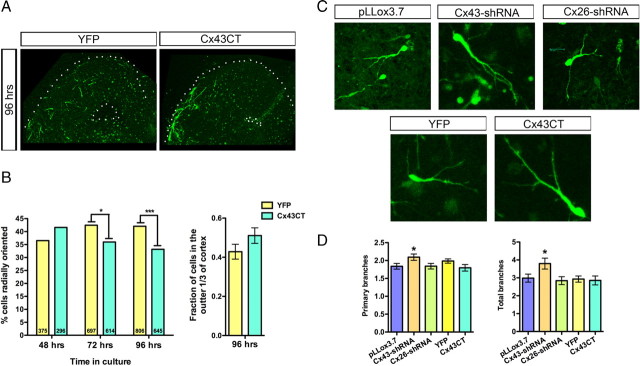
The adult cerebral cortex is composed of excitatory and inhibitory neurons that arise from progenitor cells in disparate proliferative regions in the developing brain and follow different migratory paths. Excitatory pyramidal neurons originate near the ventricle and migrate radially to their position in the cortical plate along radial glial fibers. On the other hand, inhibitory interneurons arise in the ventral telencephalon and migrate tangentially to enter the developing cortex before migrating radially to reach their correct laminar position. Gap junction adhesion has been shown to play an important mechanistic role in the radial migration of excitatory neurons. We asked whether a similar mechanism governs the tangential or radial migration of inhibitory interneurons. Using short hairpin RNA knockdown of Connexin 43 (Cx43) and Cx26 together with rescue experiments, we found that gap junctions are dispensable for the tangential migration of interneurons, but that Cx43 plays a role in the switch from tangential to radial migration that allows interneurons to enter the cortical plate and find their correct laminar position. Moreover this action is dependent on the adhesive properties and the C terminus of Cx43 but not the Cx43 channel. Thus, the radial phase of interneuron migration resembles that of excitatory neuron migration in terms of dependence on Cx43 adhesion. Furthermore, gap junctions between migrating interneurons and radial processes were observed by electron microscopy. These findings provide mechanistic and structural support for a gap junction-mediated interaction between migrating interneurons and radial glia during the switch from tangential to radial migration.
Figures
References
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Anderson SA, Marín O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
-
- Beahm DL, Oshima A, Gaietta GM, Hand GM, Smock AE, Zucker SN, Toloue MM, Chandrasekhar A, Nicholson BJ, Sosinsky GE. Mutation of a conserved threonine in the third transmembrane helix of alpha- and beta-connexins creates a dominant-negative closed gap junction channel. J Biol Chem. 2006;281:7994–8009. - PubMed
-
- Cina C, Bechberger JF, Ozog MA, Naus CC. Expression of connexins in embryonic mouse neocortical development. J Comp Neurol. 2007;504:298–313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous