

Conversion of a yeast prion protein to an infectious form in bacteria
- PMID: 20484678
- PMCID: PMC2890818
- DOI: 10.1073/pnas.0913280107
Conversion of a yeast prion protein to an infectious form in bacteria
Abstract
Prions are infectious, self-propagating protein aggregates that have been identified in evolutionarily divergent members of the eukaryotic domain of life. Nevertheless, it is not yet known whether prokaryotes can support the formation of prion aggregates. Here we demonstrate that the yeast prion protein Sup35 can access an infectious conformation in Escherichia coli cells and that formation of this material is greatly stimulated by the presence of a transplanted [PSI(+)] inducibility factor, a distinct prion that is required for Sup35 to undergo spontaneous conversion to the prion form in yeast. Our results establish that the bacterial cytoplasm can support the formation of infectious prion aggregates, providing a heterologous system in which to study prion biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
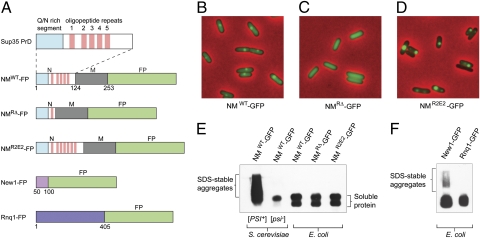
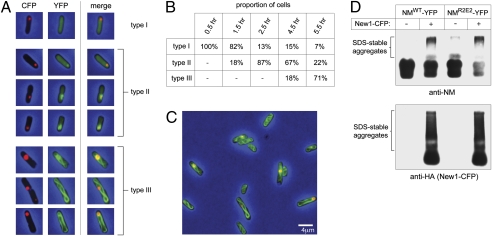
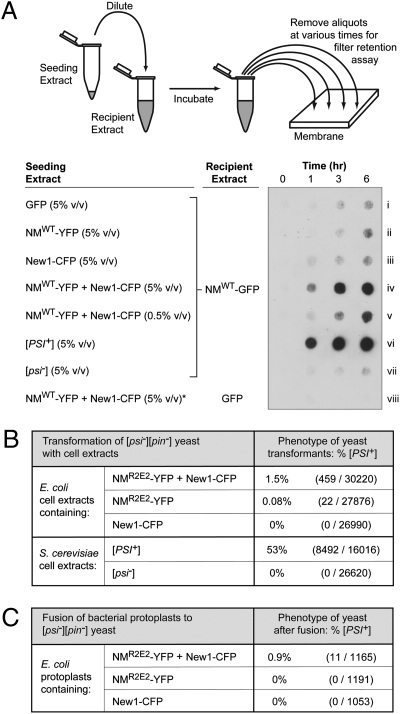
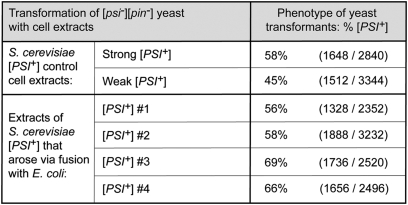
Similar articles
-
Yeast prions form infectious amyloid inclusion bodies in bacteria.Microb Cell Fact. 2012 Jun 25;11:89. doi: 10.1186/1475-2859-11-89. Microb Cell Fact. 2012. PMID: 22731490 Free PMC article.
-
Investigating the interactions of yeast prions: [SWI+], [PSI+], and [PIN+].Genetics. 2014 Jun;197(2):685-700. doi: 10.1534/genetics.114.163402. Epub 2014 Apr 11. Genetics. 2014. PMID: 24727082 Free PMC article.
-
Prion propagation can occur in a prokaryote and requires the ClpB chaperone.Elife. 2014 Aug 13;3:e02949. doi: 10.7554/eLife.02949. Elife. 2014. PMID: 25122461 Free PMC article.
-
The relationship of prions and translation.Wiley Interdiscip Rev RNA. 2010 Jul-Aug;1(1):81-9. doi: 10.1002/wrna.8. Wiley Interdiscip Rev RNA. 2010. PMID: 21339834 Free PMC article. Review.
-
The life of [PSI].Curr Genet. 2018 Feb;64(1):1-8. doi: 10.1007/s00294-017-0714-7. Epub 2017 Jun 26. Curr Genet. 2018. PMID: 28653109 Free PMC article. Review.
Cited by
-
Rebels with a cause: molecular features and physiological consequences of yeast prions.FEMS Yeast Res. 2014 Feb;14(1):136-47. doi: 10.1111/1567-1364.12116. FEMS Yeast Res. 2014. PMID: 25667942 Free PMC article. Review.
-
Yeast prions form infectious amyloid inclusion bodies in bacteria.Microb Cell Fact. 2012 Jun 25;11:89. doi: 10.1186/1475-2859-11-89. Microb Cell Fact. 2012. PMID: 22731490 Free PMC article.
-
The prion-like RNA-processing protein HNRPDL forms inherently toxic amyloid-like inclusion bodies in bacteria.Microb Cell Fact. 2015 Jul 11;14:102. doi: 10.1186/s12934-015-0284-7. Microb Cell Fact. 2015. PMID: 26160665 Free PMC article.
-
Engineered bacterial hydrophobic oligopeptide repeats in a synthetic yeast prion, [REP-PSI (+)].Front Microbiol. 2015 Apr 21;6:311. doi: 10.3389/fmicb.2015.00311. eCollection 2015. Front Microbiol. 2015. PMID: 25954252 Free PMC article.
-
The emergence of bacterial prions.PLoS Pathog. 2024 Jun 13;20(6):e1012253. doi: 10.1371/journal.ppat.1012253. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38870093 Free PMC article. Review. No abstract available.
References
-
- Aguzzi A, Polymenidou M. Mammalian prion biology: One century of evolving concepts. Cell. 2004;116:313–327. - PubMed
-
- Wickner RB, Liebman SW, Saupe SJ. Prions of yeast and filamentous fungi: [URE3+], [PSI+], [PIN+], and [Het-s] In: Prusiner SB, editor. Prion Biology and Diseases. Cold Spring Harbor Lab Press: Cold Spring Harbor, NY; 2004. pp. 305–372.
-
- Chien P, Weissman JS, DePace AH. Emerging principles of conformation-based prion inheritance. Annu Rev Biochem. 2004;73:617–656. - PubMed
-
- Kimura Y, Koitabashi S, Fujita T. Analysis of yeast prion aggregates with amyloid-staining compound in vivo. Cell Struct Funct. 2003;28:187–193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
