

The endothelial cell receptor GRP78 is required for mucormycosis pathogenesis in diabetic mice
- PMID: 20484814
- PMCID: PMC2877958
- DOI: 10.1172/JCI42164
The endothelial cell receptor GRP78 is required for mucormycosis pathogenesis in diabetic mice
Abstract
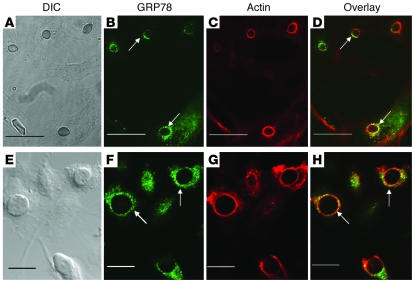
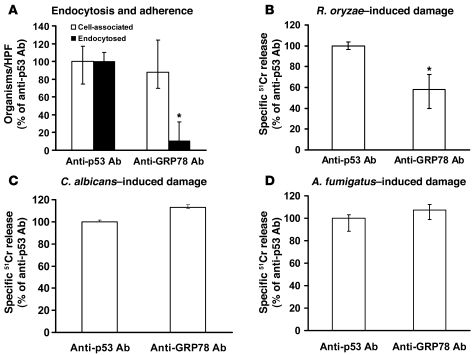
Mucormycosis is a fungal infection of the sinuses, brain, or lungs that causes a mortality rate of at least 50% despite first-line therapy. Because angioinvasion is a hallmark of mucormycosis infections, we sought to define the endothelial cell receptor(s) for fungi of the order Mucorales (the fungi that cause mucormycosis). Furthermore, since patients with elevated available serum iron, including those with diabetic ketoacidosis (DKA), are uniquely susceptible to mucormycosis, we sought to define the role of iron and glucose in regulating the expression of such a receptor. Here, we have identified glucose-regulated protein 78 (GRP78) as what we believe to be a novel host receptor that mediates invasion and damage of human endothelial cells by Rhizopus oryzae, the most common etiologic species of Mucorales, but not Candida albicans or Aspergillus fumigatus. Elevated concentrations of glucose and iron, consistent with those seen during DKA, enhanced GRP78 expression and the resulting R. oryzae invasion and damage of endothelial cells in a receptor-dependent manner. Mice with DKA, which have enhanced susceptibility to mucormycosis, exhibited increased expression of GRP78 in sinus, lungs, and brain compared with normal mice. Finally, GRP78-specific immune serum protected mice with DKA from mucormycosis. These results suggest a unique susceptibility of patients with DKA to mucormycosis and provide a foundation for the development of new therapeutic interventions for these deadly infections.
Figures








References
-
- Ibrahim AS, Edwards JE, Filler SG. Zygomycoses. In: Dismukes WE, Pappas PG, Sobel JD, eds.Clinical Mycology . New York, NY: Oxford University Press; 2003:241–251.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
