

Review
doi: 10.1038/nrm2904.
Epub 2010 May 19.
Designing customized cell signalling circuits
Affiliations
- PMID: 20485291
- PMCID: PMC2975372
- DOI: 10.1038/nrm2904
Item in Clipboard
Review
Designing customized cell signalling circuits
Nat Rev Mol Cell Biol.
2010 Jun.
Abstract
Living cells have evolved a broad array of complex signalling responses, which enables them to survive diverse environmental challenges and execute specific physiological functions. Our increasingly sophisticated understanding of the molecular mechanisms of cell signalling networks in eukaryotes has revealed a remarkably modular organization and synthetic biologists are exploring how this can be exploited to engineer cells with novel signalling behaviours. This approach is beginning to reveal the logic of how cells might evolve innovative new functions and moves us towards the exciting possibility of engineering custom cells with precise sensing-response functions that could be useful in medicine and biotechnology.
Figures
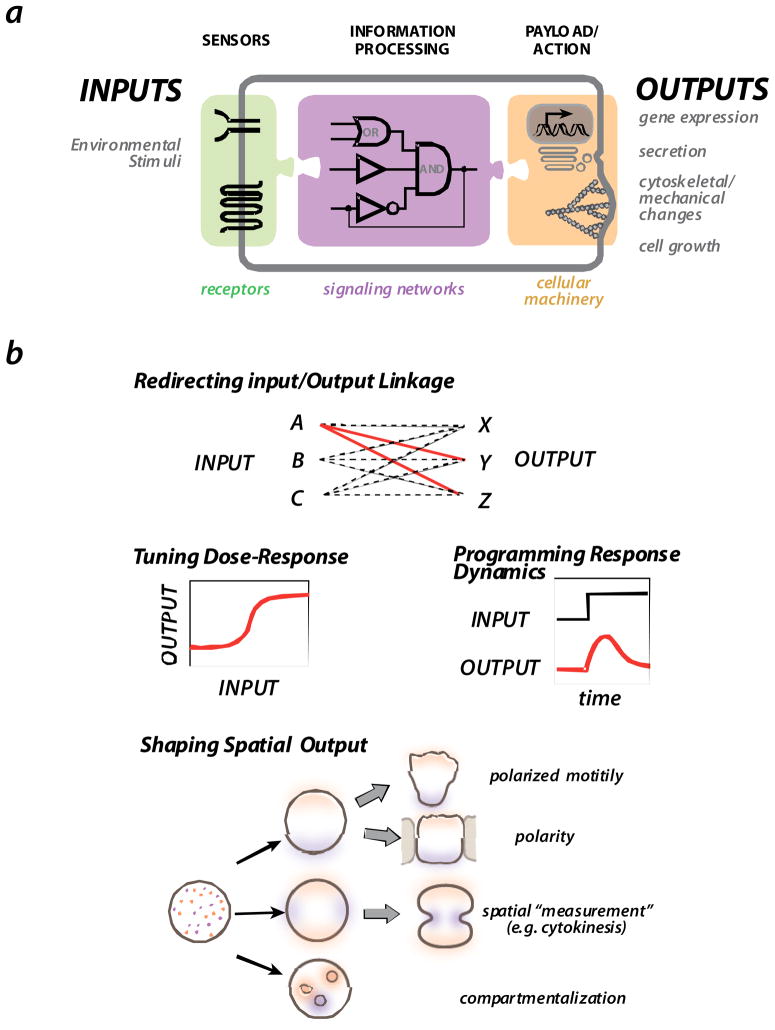
a| Cells generally sense environmental stimuli via receptors and other sensors. This information is then processed by intracellular signaling networks, which in turn engage various cellular outputs, including gene expression, secretion, cytoskeletal changes, and cell growth. b| Some of the major challenges in the evolution or engineering of novel signaling circuits are: achieving the correct linkage of specific inputs and specific outputs; tuning quantitative behaviors of the signaling response - dose-response and dynamics -- so that they are optimal for the physiological function; and generating robust spatially self-organizing processes, such as those associated with cell polarization, directed motility, cell division, and cell compartmentalization.
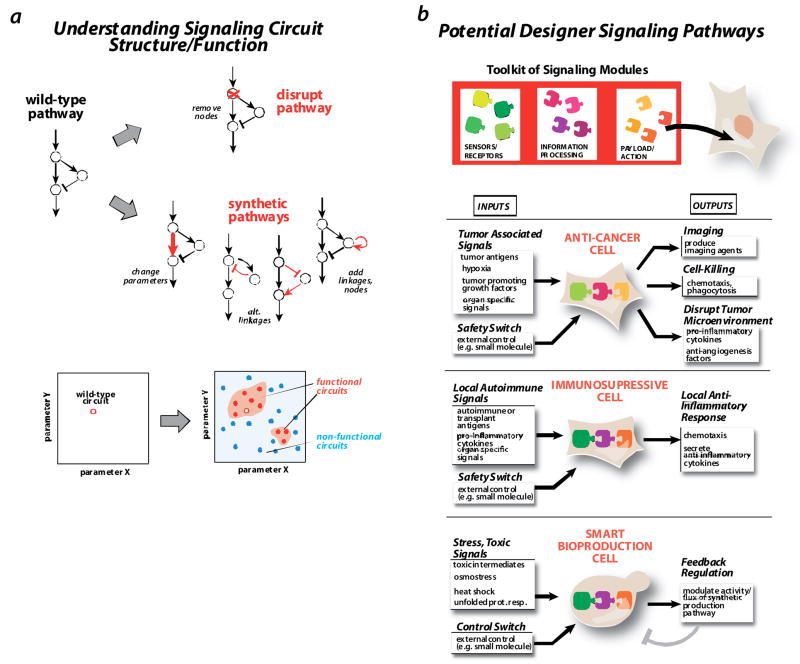
a| understanding design principles. Traditionally, methods like gene disruption are used to dissect a signaling network. Synthetic approaches offer complementary information by creating alternative versions of a network that differ both in the network connectivity or in the strength of links. By mapping the space of functional (red circles) vs. nonfunctional (blue circles) variants, one gains a deeper understanding of functional requirements. b | constructing designer signaling pathways for therapeutic or biotechnology applications. We hope to assemble a toolkit of signaling modules that can be used to create cells with designed signaling responses. An anti-cancer cell might detect a combination of tumor signals, and yield responses such as production of imaging reagents, cell killing, or secretion of factors that disrupt the tumor microenvironment. Such a cell might also have safety switches that could disable the cell if needed. An immunosuppressive cell might detect a combination of auto-immune response or transplant rejection signals, and trigger localized countermeasures, such as secretion of anti-inflammatory cytokines. A smart bioproduction (fermentation) cell would be engineered to precisely modulate the flux in growth versus production pathways in response to the stress state of the cell, thus optimizing overall yield.
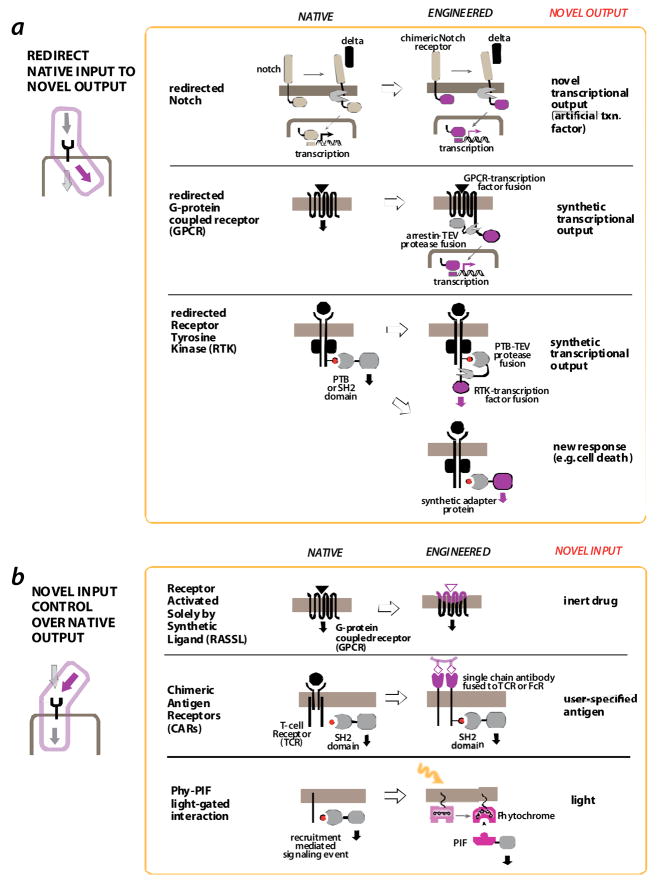
a | redirecting native inputs to novel outputs. The C-terminal domain of the notch receptor is transcription factor that is released by transmembrane proteolysis upon activation by the ligand, delta. Replacement with an alternative transcription factor domain yields a new gene expression response [26]. G-protein coupled receptor (GPCR) output can be redirected in a similar way by fusing a transcription factor domain via a tether with a TEV protease site. Activation of the GPCR results in recruitment of the protein arrestin. If an arrestin-TEV protease fusion is expressed in the cell, GPCR activation results in release of the transcription factor, and a novel gene expression output [28]. Thus GPCR activation can be arbitrarily linked to a novel transcriptional output. Receptor tyrosine kinase (RTK) output can be redirected by harnessing the recruitment of synthetic SH2 or PTB domain adapters to the activated and tyrosine phosphorylated receptor. The SH2 domain could be used to recruit a TEV protease (to again release an artificially tethered transcriptional domain) [28] or to recruit novel effector domains, such as those involved in cell death [29]. b | engineering novel input control over native responses. GPCRs have been engineered to be controlled by small molecule agonists by mutating their extracellular surface such that they no longer bind their endogenous ligands (Receptors activated solely by sythetic ligand – RASSL [32]). Receptors that activate T-cells in response to arbitrary inputs can be generated by fusing engineered single chain antibodies (scFv’s) to the intracellular region of the T-cell receptor (CD3 zeta chain) – referred to as chimeric antigen receptors (CARs [16, 17]). A recruitment mediated signaling event can be placed under light control by replacing the endogenous interaction with the light-gated interaction Phytochrome-PIF interaction pair from plants [44].
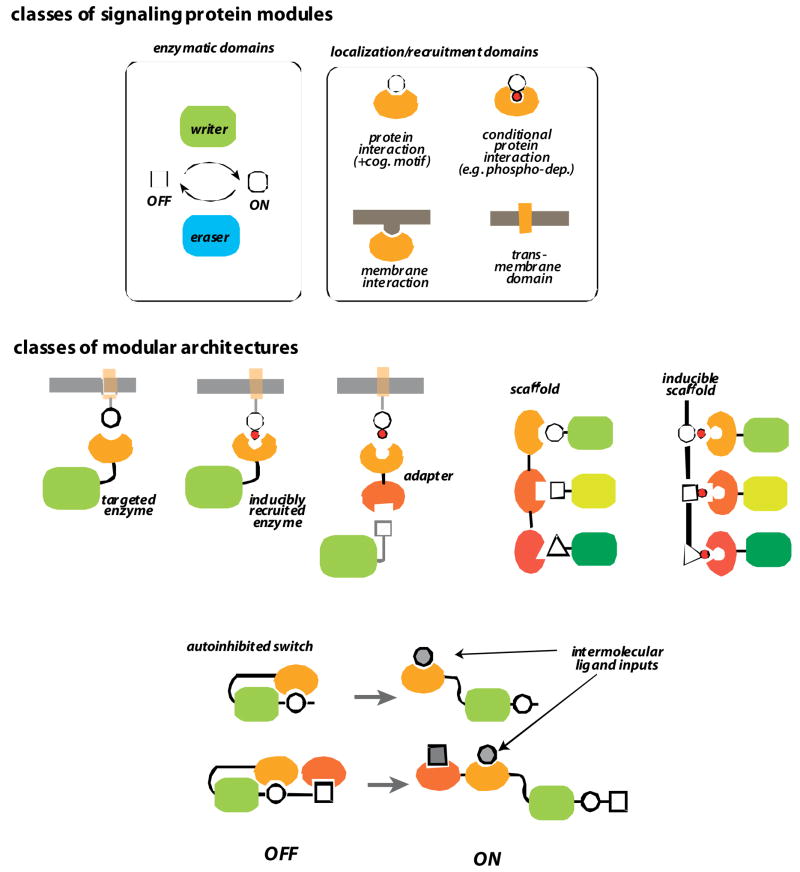
a | enzymatic and regulatory domains. Modular eukaryotic signaling proteins are generally composed of enzymatic domains and localization domains. Enzymatic domains, like kinases and phosphatases, and GEFs and GAPs, catalyze regulatory modifications such as phosphorylation and GTPase activation, respectively (enzymatic domains often come in “writer” and “eraser” pairs that have opposing activities). These enzymatic domains are regulated and targeted by interaction domains, including protein-protein interaction domains, membrane interaction domains, or transmembrane domains. b | different classes of multidomain architectures. Enzymatic domains can be directly targeted to specific substrates, partners or subcellular locations by interaction domains. Alternatively they can be indirectly targeted via adapters or scaffold proteins, which contain multiple interaction domains. Interaction domains can also allosterically regulate catalytic domains by engaging in intramolecular autoinhibitory interactions. Such switch proteins can be activated by competing ligands that relieve autoinhibition.
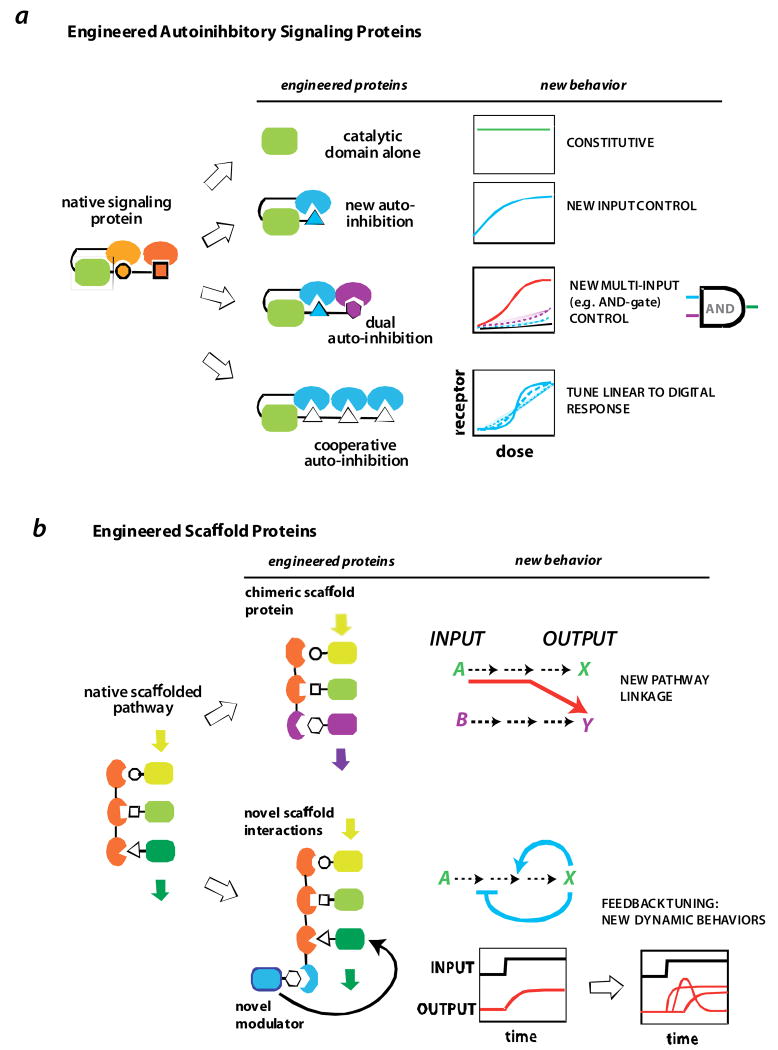
a | engineered allosteric protein switches. Dueber et al [49,51] showed that the allosterically regulation of the signaling protein N-WASP could be reprogrammed by recombining the catalytic domain from N-WASP with different combinations of interaction domains. Novel behaviors included multi-input (AND-gate) control and highly cooperative switch-like activation. b | using scaffold proteins as a molecular circuitboard for reshaping signaling output. The input/output linkage of a MAP kinase pathway in yeast could be redirected via an engineered chimeric scaffold that assembled a novel combination of kinases [58]. Novel interaction sites can also be appended on to scaffolds to recruit additional modulatory factors. These additional factors can be build synthetic feedback loops that can be used to generate pathways that display diverse signaling dynamics [62].
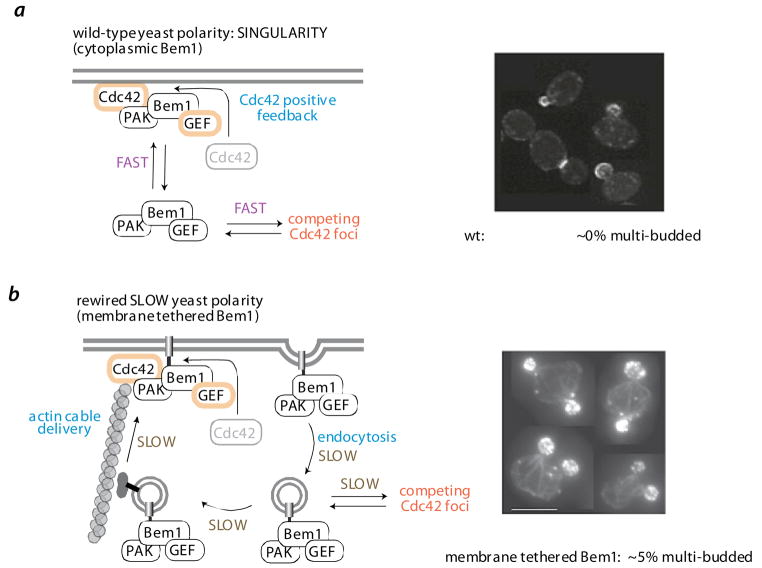
a | wild-type polarization circuit controls single bud formation. In budding yeast, localized activation of the polarity GTPase Cdc42 is amplified by a positive feedback loop - active Cdc42 recruits the scaffold protein Bem1, co-assembles the p21 activated kinase (PAK – Ste20) and the Cdc42 GEF (Cdc24). Although a cell may have multiple Cdc42 foci, these are quickly resolved into one dominant foci, which develops into the cells only bud. A fast rate of interchange of the diffusible Bem1/PAK/GEF complex between competing Cdc42 foci, is hypothesized to be critical for resolution into a single dominant foci. b | a synthetic slow polarization circuit leads to multiple bud formation. To test this hypothesis, Bem1 was artificially tethered to the membrane via a fused membrane targeting motif [65]. Although this membrane tethered Bem1 can properly assemble the Bem1/PAK/GEF complex at sites of Cdc42 activity (i.e. the positive feedback loop), the exchange of the complex between competing Cdc42 foci is slow (dependent on vesicular transport via actin cables and endocytosis). This synthetic polarization circuit therefore leads to poor resolution of competing Cdc42 foci and a much higher frequency (5% vs ~ 0%) of multibudded cells (micrographs from [65]).
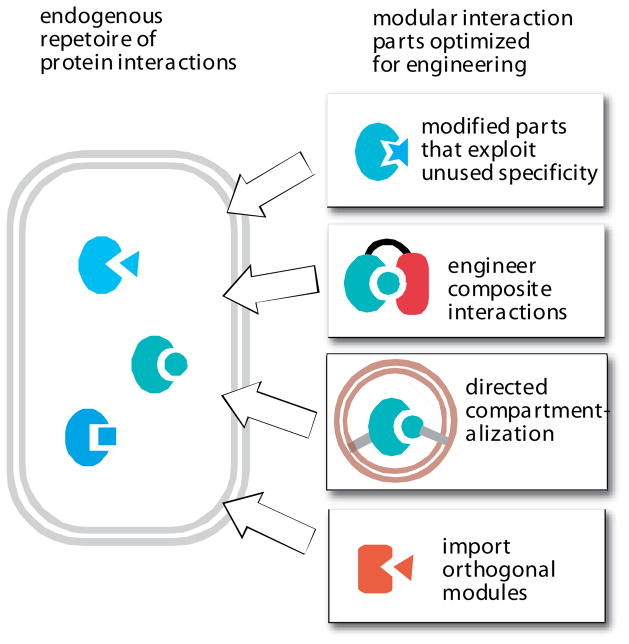
A native cell has its own native repetoire of protein interaction modules, and thus it is challenging to engineer new functions using related interaction modules that might show inadvertent and unintended crosstalk in the cell. An optimized toolkit of interaction parts could significantly increase the predictability of cellular engineering, by eliminating the chance for unintended crosstalk. Several strategies for optimization include engineering of interaction modules that exploit untapped specificity; engineering of composite, multi-domain interactions; combining interaction modules with novel subcellular targeting; and importing orthogonal interaction modules (either synthetically constructed or from other organisms) that are not found in the host cell.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
