

Myosin II contributes to cell-scale actin network treadmilling through network disassembly
- PMID: 20485438
- PMCID: PMC3662466
- DOI: 10.1038/nature08994
Myosin II contributes to cell-scale actin network treadmilling through network disassembly
Abstract
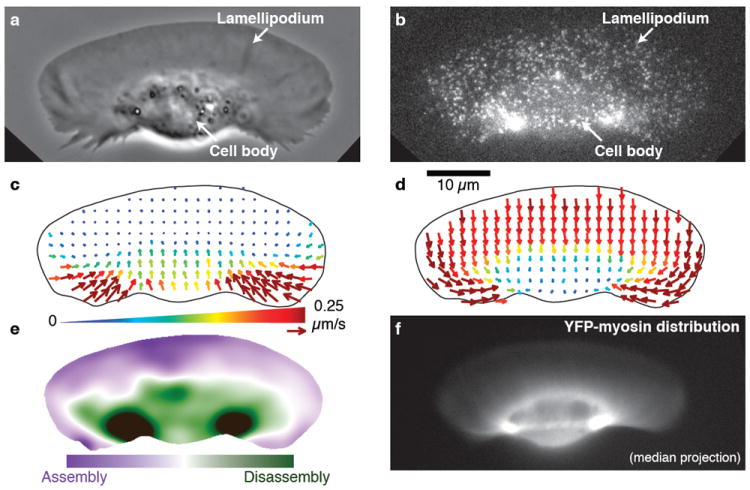
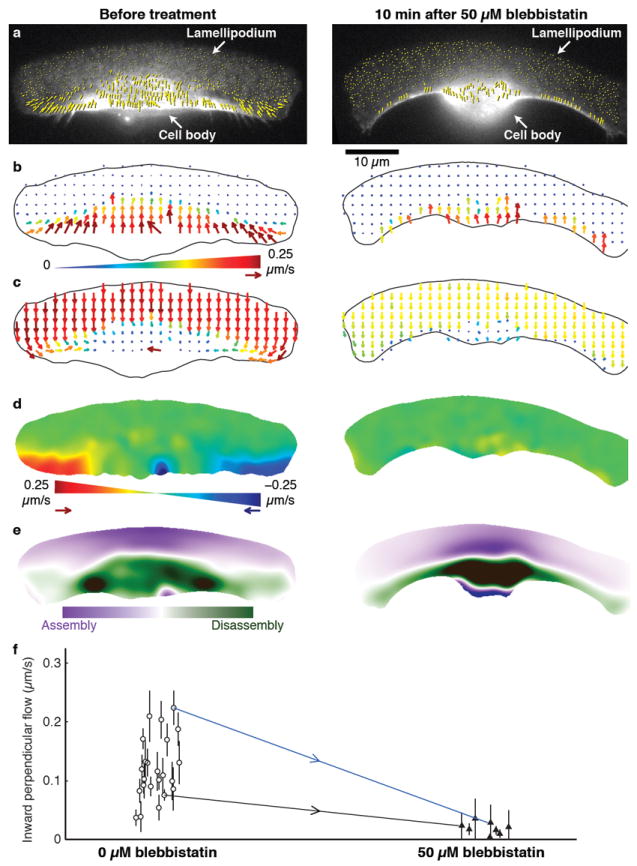
Crawling locomotion of eukaryotic cells is achieved by a process dependent on the actin cytoskeleton: protrusion of the leading edge requires assembly of a network of actin filaments, which must be disassembled at the cell rear for sustained motility. Although ADF/cofilin proteins have been shown to contribute to actin disassembly, it is not clear how activity of these locally acting proteins could be coordinated over the distance scale of the whole cell. Here we show that non-muscle myosin II has a direct role in actin network disassembly in crawling cells. In fish keratocytes undergoing motility, myosin II is concentrated in regions at the rear with high rates of network disassembly. Activation of myosin II by ATP in detergent-extracted cytoskeletons results in rear-localized disassembly of the actin network. Inhibition of myosin II activity and stabilization of actin filaments synergistically impede cell motility, suggesting the existence of two disassembly pathways, one of which requires myosin II activity. Our results establish the importance of myosin II as an enzyme for actin network disassembly; we propose that gradual formation and reorganization of an actomyosin network provides an intrinsic destruction timer, enabling long-range coordination of actin network treadmilling in motile cells.
Figures


References
-
- Abercrombie M. The Croonian lecture, 1978: the crawling movement of metazoan cells. Proc R Soc Lond B Biol Sci. 1980;207:129–47.
-
- Theriot JA, Mitchison TJ. Actin microfilament dynamics in locomoting cells. Nature. 1991;352:126–31. - PubMed
-
- Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–65. - PubMed
-
- Euteneuer U, Schliwa M. Persistent, directional motility of cells and cytoplasmic fragments in the absence of microtubules. Nature. 1984;310:58–61. - PubMed
-
- Small JV, Herzog M, Häner M, Abei U. Visualization of actin filaments in keratocyte lamellipodia: negative staining compared with freeze-drying. J Struct Biol. 1994;113:135–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
