

The alternative oxidase (AOX) gene in Vibrio fischeri is controlled by NsrR and upregulated in response to nitric oxide
- PMID: 20487270
- PMCID: PMC3682826
- DOI: 10.1111/j.1365-2958.2010.07194.x
The alternative oxidase (AOX) gene in Vibrio fischeri is controlled by NsrR and upregulated in response to nitric oxide
Abstract
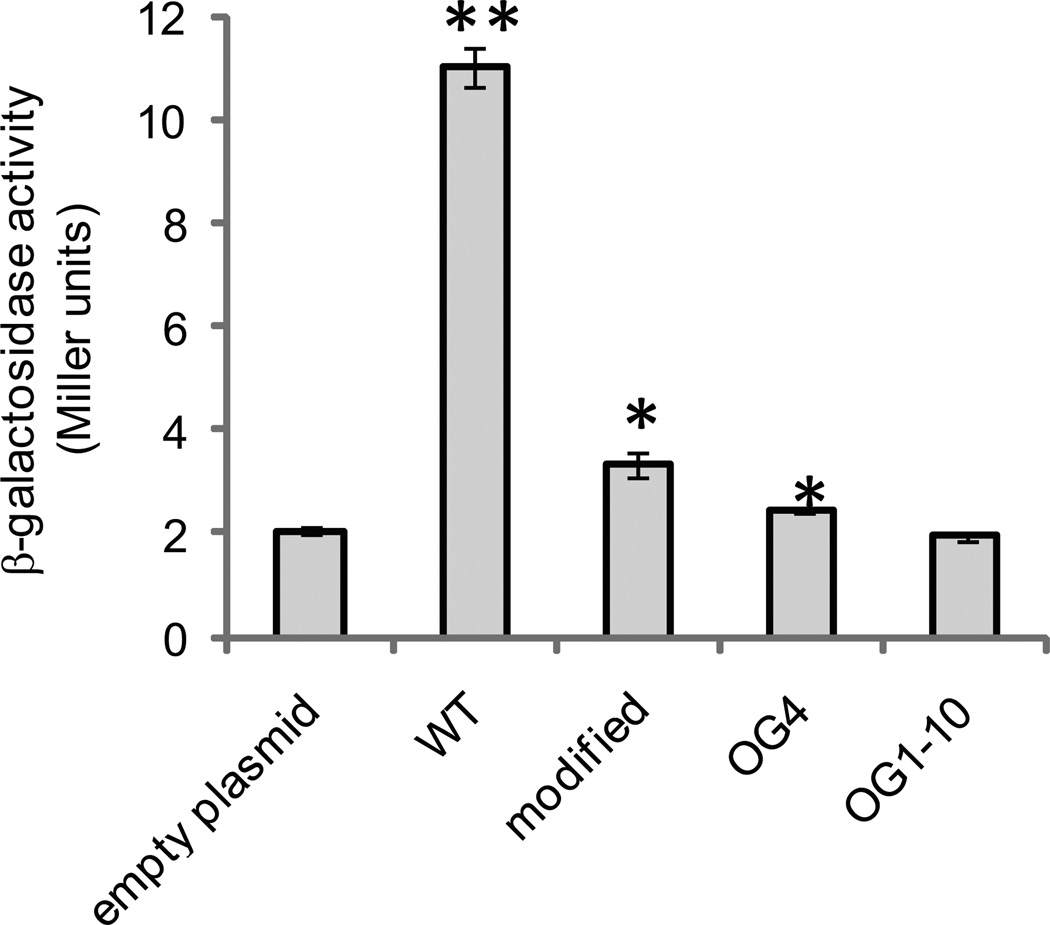
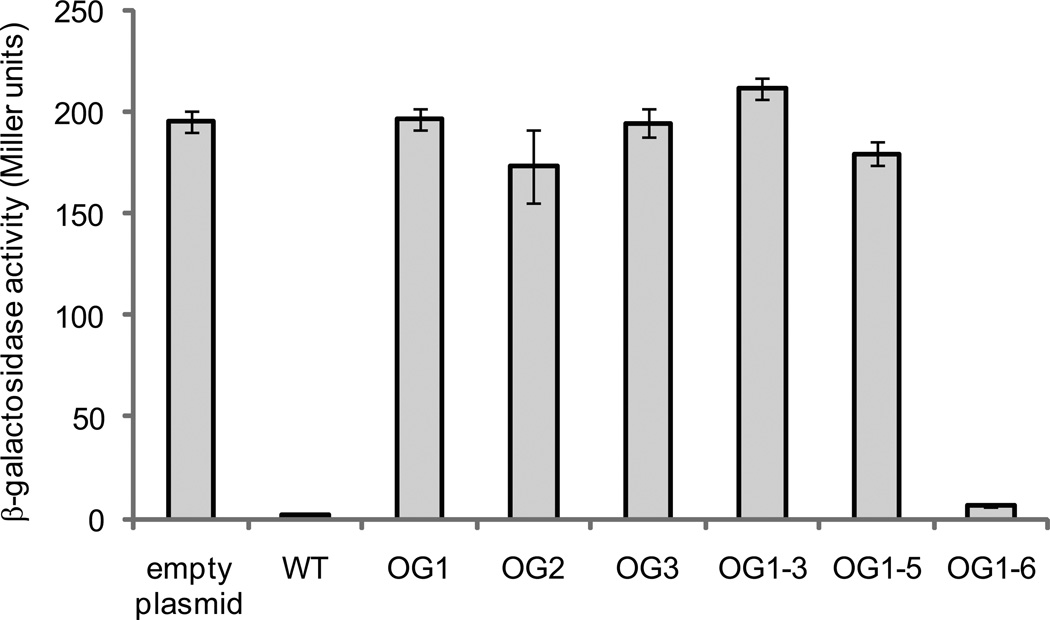
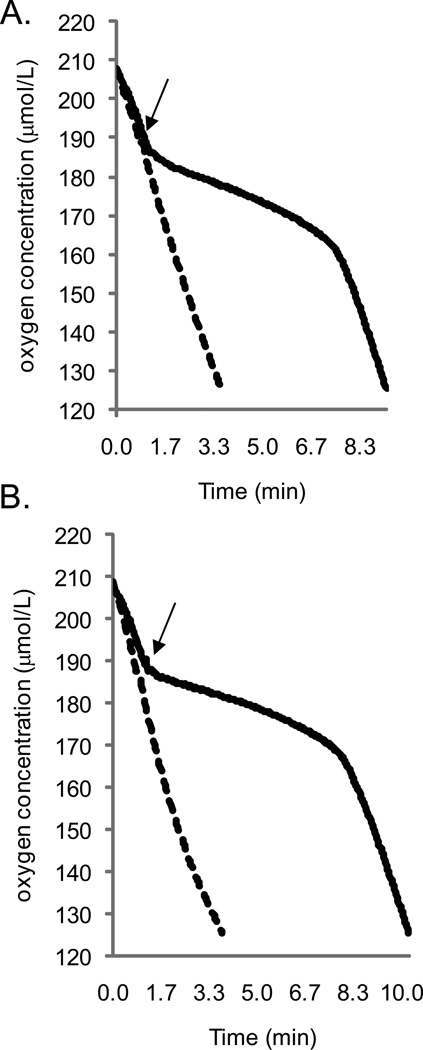
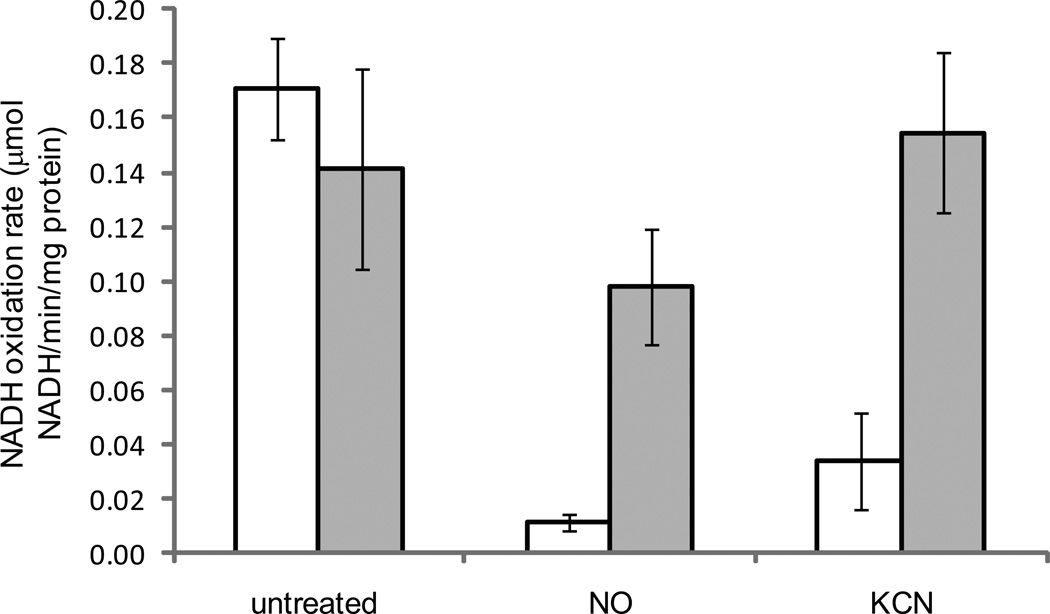
Alternative oxidase (AOX) is a respiratory oxidase found in certain eukaryotes and bacteria; however, its role in bacterial physiology is unclear. Exploiting the genetic tractability of the bacterium Vibrio fischeri, we explore the regulation of aox expression and AOX function. Using quantitative PCR and reporter assays, we demonstrate that aox expression is induced in the presence of nitric oxide (NO), and that the NO-responsive regulatory protein NsrR mediates the response. We have identified key amino acid residues important for NsrR function and experimentally confirmed a bioinformatically predicted NsrR binding site upstream of aox. Microrespirometry demonstrated that oxygen consumption by V. fischeri CydAB quinol oxidase is inhibited by NO treatment, whereas oxygen consumption by AOX is less sensitive to NO. NADH oxidation assays using inverted membrane vesicles confirmed that NO directly inhibits CydAB, and that AOX is resistant to NO inhibition. These results indicate a role for V. fischeri AOX in aerobic respiration during NO stress.
Figures

Comment in
-
An alternative route to nitric oxide resistance.Mol Microbiol. 2010 Jul 1;77(1):6-10. doi: 10.1111/j.1365-2958.2010.07195.x. Epub 2010 May 4. Mol Microbiol. 2010. PMID: 20487269 Review.
References
-
- Berthold DA, Voevodskaya N, Stenmark P, Graslund A, Nordlund P. EPR studies of the mitochondrial alternative oxidase. Evidence for a diiron carboxylate center. J. Biol. Chem. 2002;277:43608–43614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
