

The leader intron of AtMHX can elicit, in the absence of splicing, low-level intron-mediated enhancement that depends on the internal intron sequence
- PMID: 20487561
- PMCID: PMC3095359
- DOI: 10.1186/1471-2229-10-93
The leader intron of AtMHX can elicit, in the absence of splicing, low-level intron-mediated enhancement that depends on the internal intron sequence
Abstract
Background: Introns stimulate gene expression in a wide range of organisms by increasing the levels of mature mRNA, without affecting mRNA stability. Although introns sometimes function as transcriptional enhancers, they usually stimulate expression by a process termed intron-mediated enhancement (IME). The mechanism of IME is largely unknown. While splicing per se is not sufficient for IME, as evident from the fact that not all introns increase expression, it is not clear yet whether splicing of the enhancing introns is essential for enhancement. The leader intron (LI) of the Arabidopsis AtMHX gene was previously shown to substantially increase the expression of the AtMHX promoter. Here we investigated whether this LI acts as a transcriptional enhancer and whether its splicing is essential for IME.
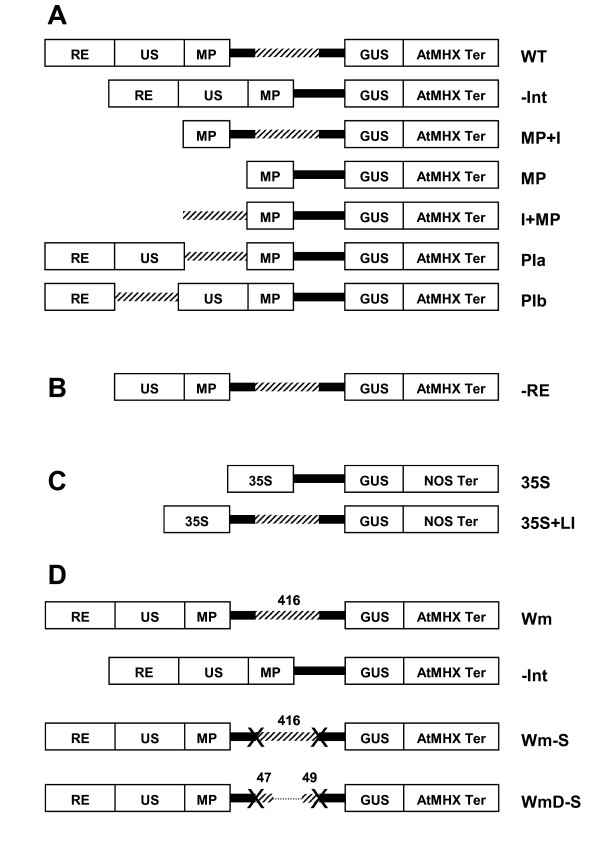
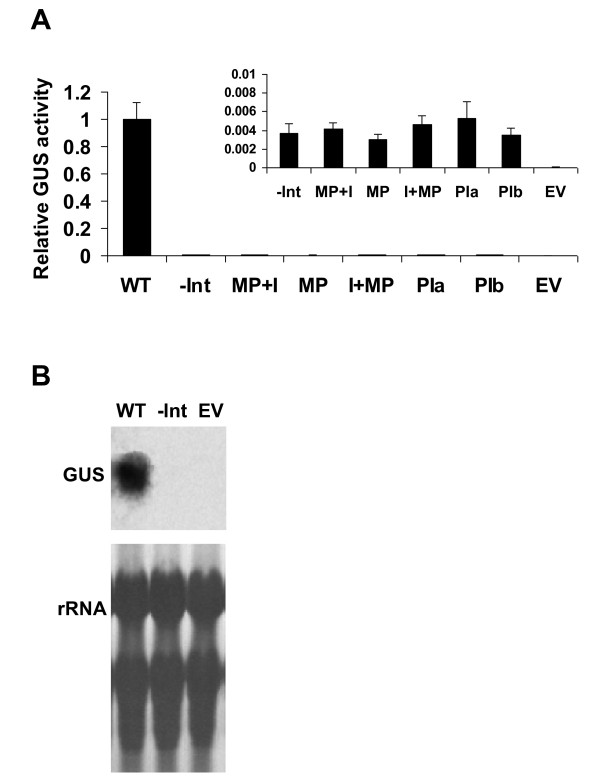
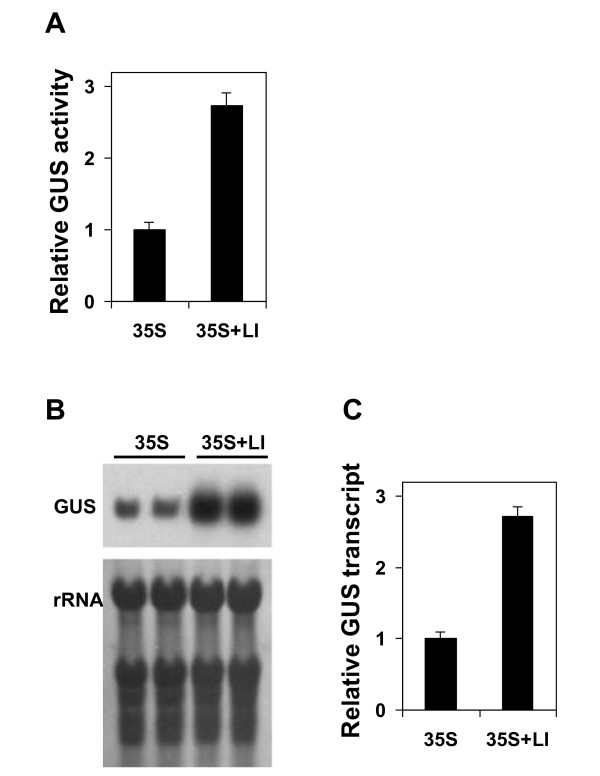
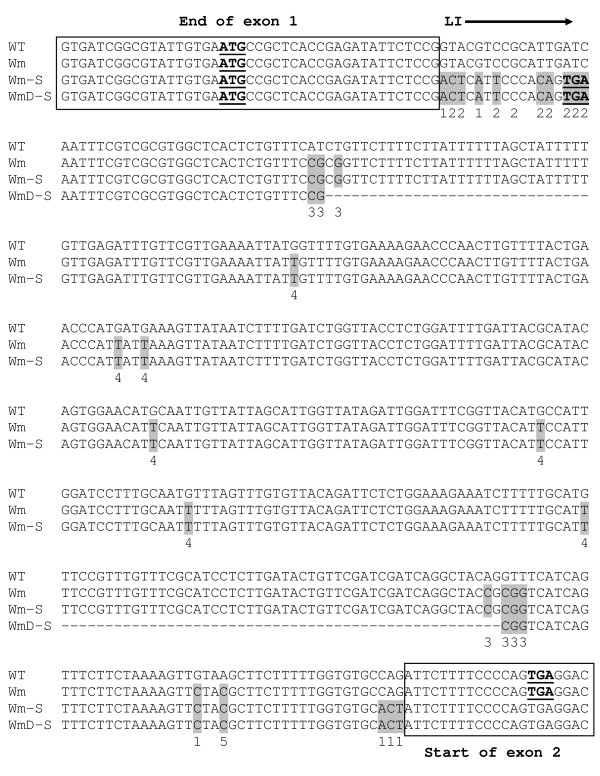
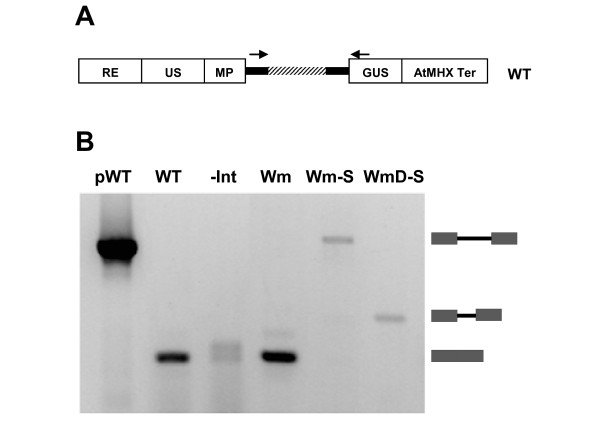
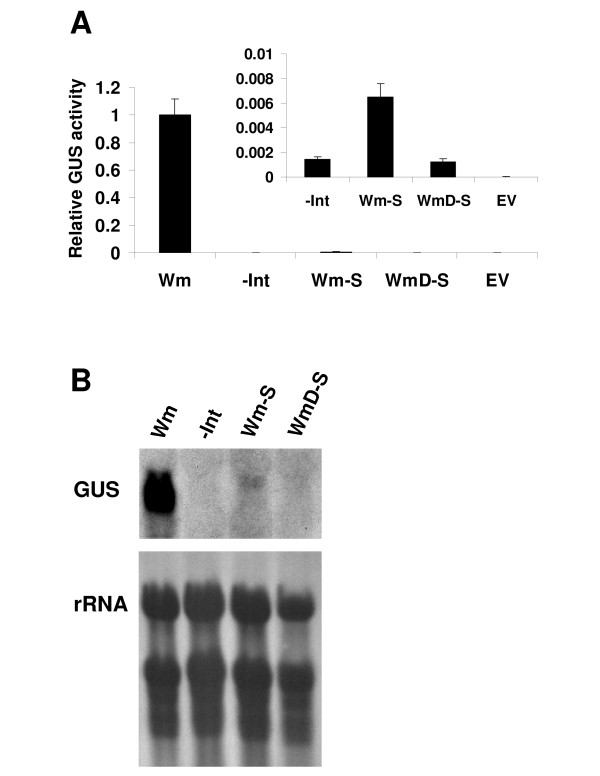
Results: Expression in transformed Arabidopsis plants of an AtMHX::GUS construct from which the LI was eliminated was similar to a construct that included only the minimal promoter fused to GUS. Yet, almost no expression was seen in constructs that included the LI in addition to the minimal promoter or the LI inserted in various locations in the promoter. While the LI enhanced 272-fold the expression of the weak AtMHX promoter, only a 3-fold enhancement was observed for the strong CaMV 35S promoter. In the context of the AtMHX promoter, an unspliceable version of the LI that had mutated 5' and 3' splice sites mediated a low-level (5-fold) enhancement. Eliminating the internal 320 nt of the 416 nt unspliceable intron resulted in loss of ability to mediate low-level enhancement.
Conclusions: Although AtMHX promoter shows almost no expression in the absence of its LI, this intron does not act as a transcriptional enhancer and is unable to support expression in the absence of the enhancer elements of the promoter. It is also shown that the same intron can have very different contributions to expression of different promoters. Our results also demonstrate that while splicing is essential for substantial IME, in the absence of splicing low-level enhancement can be obtained. Notably, it is shown that the internal intron sequence plays a significant role in mediating the low-level enhancement of unspliced introns.
Figures
References
-
- Rose AB. In: Current Topics in Microbiology and Immunology. Reddy ASN, Golovkin M, editor. Vol. 326. Berlin Heidelberg, Springer-Verlag; 2008. Rose, A.B. (2008) Intron-mediated regulation of gene expression; pp. 277–290. full_text. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
