

CNS-expressed cathepsin D prevents lymphopenia in a murine model of congenital neuronal ceroid lipofuscinosis
- PMID: 20489146
- PMCID: PMC2893670
- DOI: 10.2353/ajpath.2010.091267
CNS-expressed cathepsin D prevents lymphopenia in a murine model of congenital neuronal ceroid lipofuscinosis
Abstract
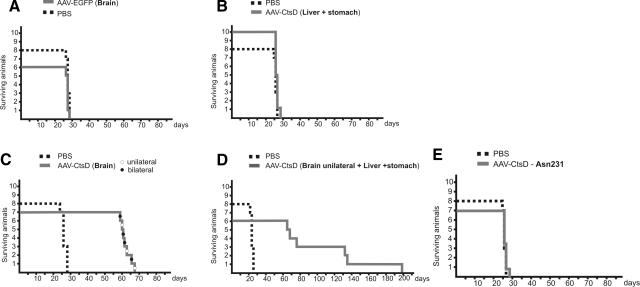
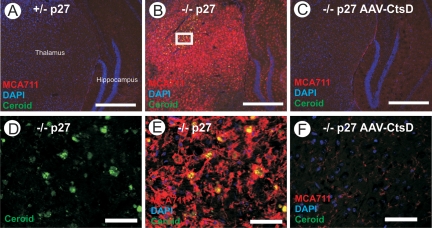
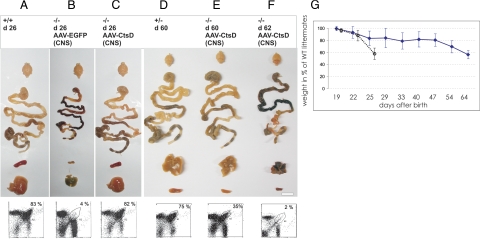
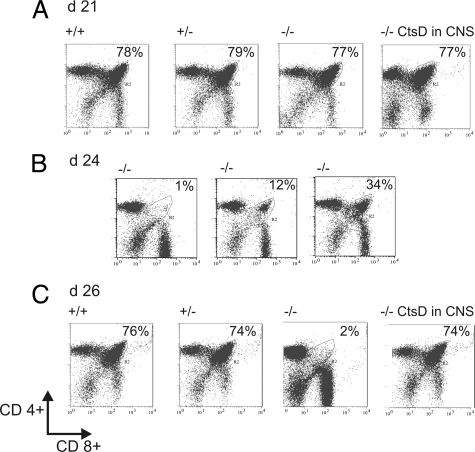
Deficiency in Cathepsin D (CtsD), the major cellular lysosomal aspartic proteinase, causes the congenital form of neuronal ceroid lipofuscinoses (NCLs). CtsD-deficient mice show severe visceral lesions like lymphopenia in addition to their central nervous system (CNS) phenotype of ceroid accumulation, microglia activation, and seizures. Here we demonstrate that re-expression of CtsD within the CNS but not re-expression of CtsD in visceral organs prevented both central and visceral pathologies of CtsD(-/-) mice. Our results suggest that CtsD was substantially secreted from CNS neurons and drained from CNS to periphery via lymphatic routes. Through this drainage, CNS-expressed CtsD acts as an important modulator of immune system maintenance and peripheral tissue homeostasis. These effects depended on enzymatic activity and not on proposed functions of CtsD as an extracellular ligand. Our results furthermore demonstrate that the prominent accumulation of ceroid/lipofuscin and activation of microglia in brains of CtsD(-/-) are not lethal factors but can be tolerated by the rodent CNS.
Figures
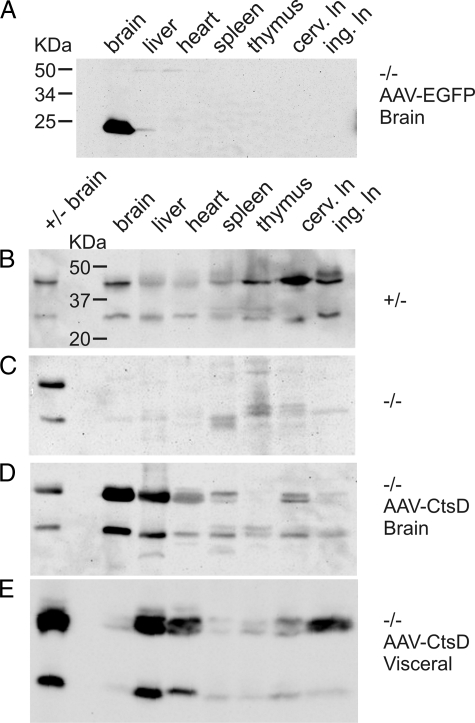
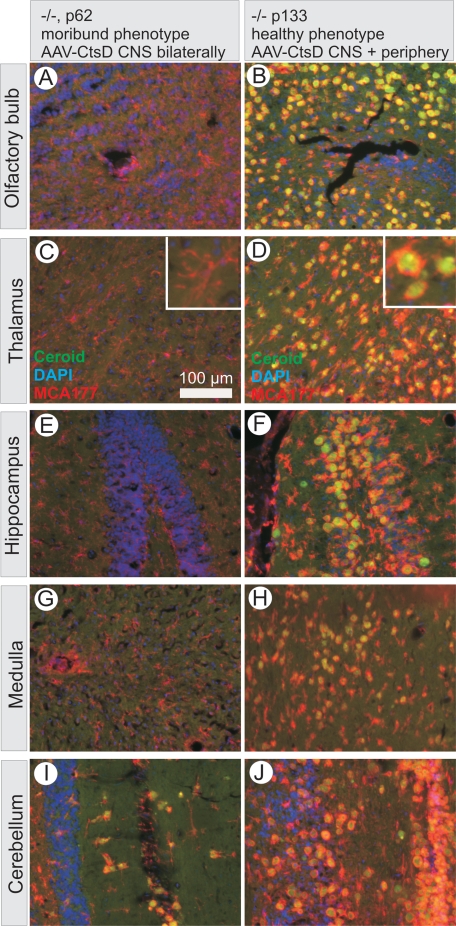
Similar articles
-
Enzyme replacement therapy with recombinant pro-CTSD (cathepsin D) corrects defective proteolysis and autophagy in neuronal ceroid lipofuscinosis.Autophagy. 2020 May;16(5):811-825. doi: 10.1080/15548627.2019.1637200. Epub 2019 Jul 16. Autophagy. 2020. PMID: 31282275 Free PMC article.
-
Lack of Cathepsin D in the central nervous system results in microglia and astrocyte activation and the accumulation of proteinopathy-related proteins.Sci Rep. 2022 Jul 8;12(1):11662. doi: 10.1038/s41598-022-15805-3. Sci Rep. 2022. PMID: 35804072 Free PMC article.
-
Neuroectoderm-specific deletion of cathepsin D in mice models human inherited neuronal ceroid lipofuscinosis type 10.Biochimie. 2016 Mar;122:219-26. doi: 10.1016/j.biochi.2015.07.020. Epub 2015 Jul 29. Biochimie. 2016. PMID: 26232697
-
Developmental changes in the expression of neuronal ceroid lipofuscinoses-linked proteins.Mol Genet Metab. 2000 Sep-Oct;71(1-2):190-4. doi: 10.1006/mgme.2000.3071. Mol Genet Metab. 2000. PMID: 11001810 Review.
-
A Novel Variant of the CTSD Gene Associated with Juvenile-onset Neuronal Ceroid Lipofuscinosis Type 10: A Case Report and Literature Review.Cerebellum. 2024 Dec 10;24(1):7. doi: 10.1007/s12311-024-01773-z. Cerebellum. 2024. PMID: 39656415 Review.
Cited by
-
Advances in the Treatment of Neuronal Ceroid Lipofuscinosis.Expert Opin Orphan Drugs. 2019;7(11):473-500. doi: 10.1080/21678707.2019.1684258. Epub 2019 Nov 27. Expert Opin Orphan Drugs. 2019. PMID: 33365208 Free PMC article.
-
Overexpression of BDNF increases excitability of the lumbar spinal network and leads to robust early locomotor recovery in completely spinalized rats.PLoS One. 2014 Feb 14;9(2):e88833. doi: 10.1371/journal.pone.0088833. eCollection 2014. PLoS One. 2014. PMID: 24551172 Free PMC article.
-
Current and Emerging Treatment Strategies for Neuronal Ceroid Lipofuscinoses.CNS Drugs. 2019 Apr;33(4):315-325. doi: 10.1007/s40263-019-00620-8. CNS Drugs. 2019. PMID: 30877620 Free PMC article. Review.
-
Ocular therapies for neuronal ceroid lipofuscinoses: more than meets the eye.Neural Regen Res. 2022 Aug;17(8):1755-1756. doi: 10.4103/1673-5374.332148. Neural Regen Res. 2022. PMID: 35017433 Free PMC article. No abstract available.
-
Long-Term Assessment of AAV-Mediated Zinc Finger Nuclease Expression in the Mouse Brain.Front Mol Neurosci. 2017 May 23;10:142. doi: 10.3389/fnmol.2017.00142. eCollection 2017. Front Mol Neurosci. 2017. PMID: 28588449 Free PMC article.
References
-
- Baechle D, Flad T, Cansier A, Steffen H, Schittek B, Tolson J, Herrmann T, Dihazi H, Beck A, Mueller GA, Mueller M, Stevanovic S, Garbe C, Mueller CA, Kalbacher H. Cathepsin D is present in human eccrine sweat and involved in the postsecretory processing of the antimicrobial peptide DCD-1L. J Biol Chem. 2006;281:5406–5415. - PubMed
-
- Saftig P, Peters C, von Figura K, Craessaerts K, Van Leuven F, De Strooper B. Amyloidogenic processing of human amyloid precursor protein in hippocampal neurons devoid of cathepsin D. J Biol Chem. 1996;271:27241–27244. - PubMed
-
- Lkhider M, Castino R, Bouguyon E, Isidoro C, Ollivier-Bousquet M. Cathepsin D: released by lactating rat mammary epithelial cells is involved in prolactin cleavage under physiological conditions. J Cell Sci. 2004;117:5155–5164. - PubMed
-
- Hilfiker-Kleiner D, Kaminski K, Podewski E, Bonda T, Schaefer A, Sliwa K, Forster O, Quint A, Landmesser U, Doerries C, Luchtefeld M, Poli V, Schneider MD, Balligand JL, Desjardins F, Ansari A, Struman I, Nguyen NQ, Zschemisch NH, Klein G, Heusch G, Schulz R, Hilfiker A, Drexler H. A cathepsin D-cleaved 16 kDa form of prolactin mediates postpartum cardiomyopathy. Cell. 2007;128:589–600. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
