

Pheromones in birds: myth or reality?
- PMID: 20490809
- PMCID: PMC3522863
- DOI: 10.1007/s00359-010-0534-4
Pheromones in birds: myth or reality?
Abstract
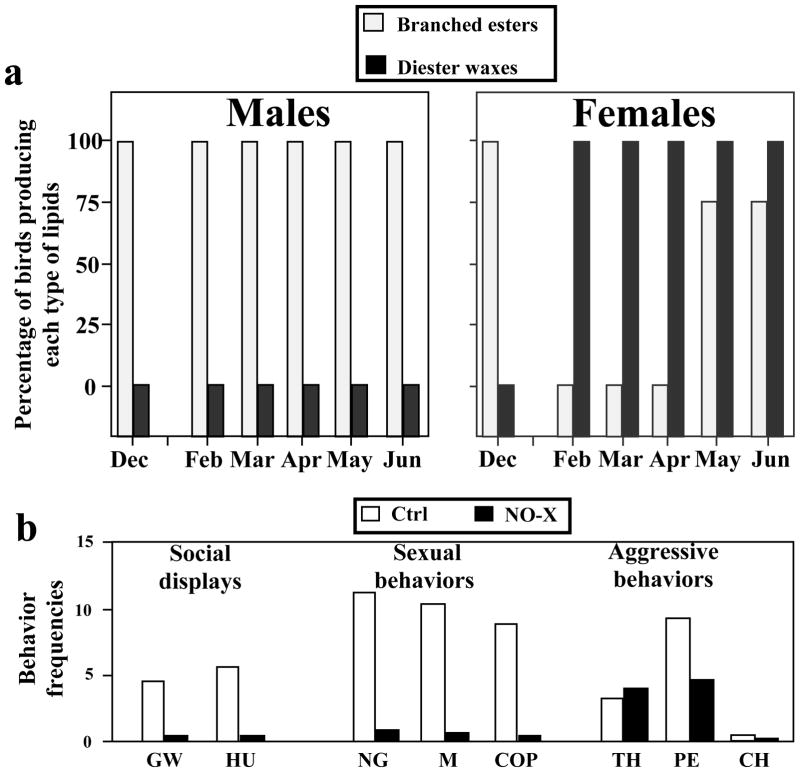
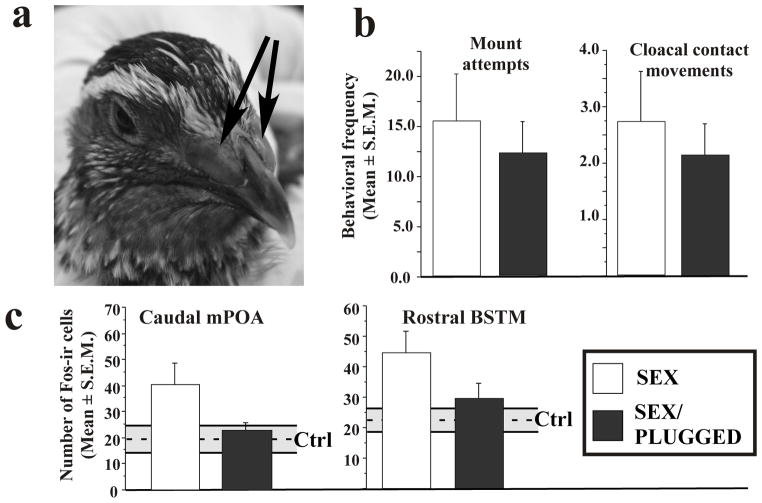
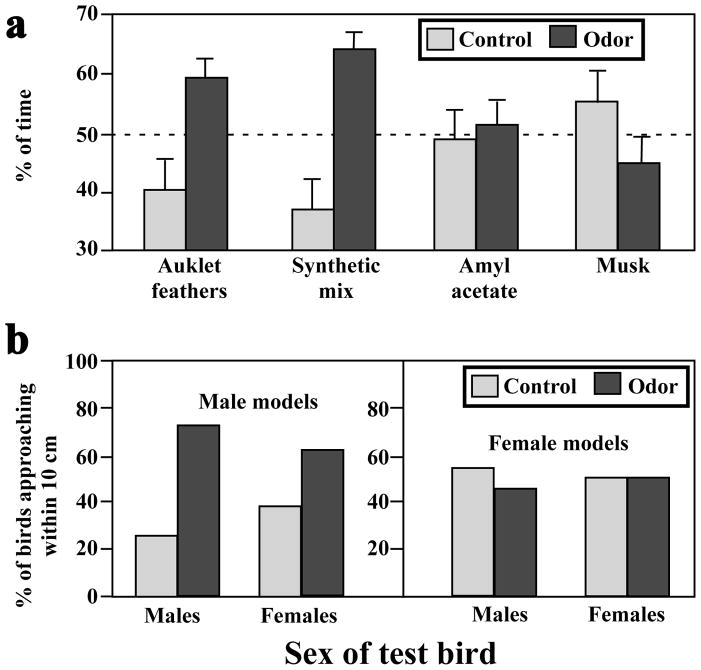
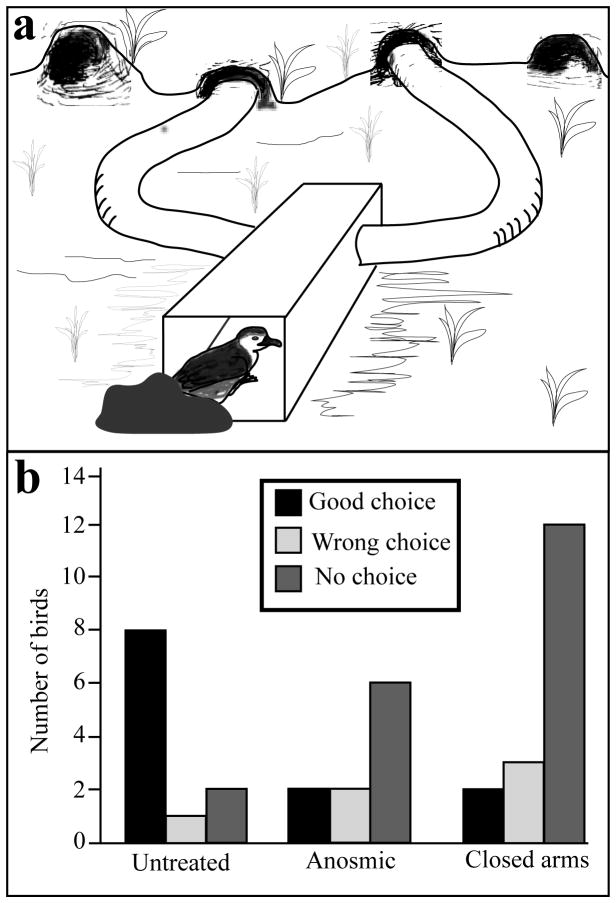
Birds are anosmic or at best microsmatic… This misbelief persisted until very recently and has strongly influenced the outcome of communication studies in birds, with olfaction remaining neglected as compared to acoustic and visual channels. However, there is now clear empirical evidence showing that olfaction is perfectly functional in birds and birds use olfactory information in a variety of ethological contexts. Although the existence of pheromones has never been formally demonstrated in this vertebrate class, different groups of birds, such as petrels, auklets and ducks have been shown to produce specific scents that could play a significant role in within-species social interactions. Behavioral experiments have indeed demonstrated that these odors influence the behavior of conspecifics. Additionally, in quail, deprivation of olfactory inputs decreases neuronal activation induced by sexual interactions with a female. It seems therefore well established that birds enjoy a functional sense of smell and a fast growing body of experimental evidence suggests that they use this channel of olfactory communication to control their social life. The unequivocal identification of an avian pheromone is, however, still ahead of us but there are now many exciting opportunities to unravel the behavioral and physiological particularities of chemical communication in birds.
Figures
Similar articles
-
The perfume of reproduction in birds: chemosignaling in avian social life.Horm Behav. 2015 Feb;68:25-42. doi: 10.1016/j.yhbeh.2014.06.001. Epub 2014 Jun 11. Horm Behav. 2015. PMID: 24928570 Free PMC article. Review.
-
Biological responsiveness to pheromones provides fundamental and unique insight into olfactory function.Chem Senses. 1996 Apr;21(2):245-56. doi: 10.1093/chemse/21.2.245. Chem Senses. 1996. PMID: 8670703 Review.
-
Species, gender, and identity: cracking petrels' sociochemical code.Chem Senses. 2010 May;35(4):309-21. doi: 10.1093/chemse/bjq021. Epub 2010 Feb 26. Chem Senses. 2010. PMID: 20190009
-
Chemical communication in invertebrates.Experientia. 1986 Mar 15;42(3):232-41. doi: 10.1007/BF01942503. Experientia. 1986. PMID: 3514260 Review. No abstract available.
-
Large-scale characterization of sex pheromone communication systems in Drosophila.Nat Commun. 2021 Jul 6;12(1):4165. doi: 10.1038/s41467-021-24395-z. Nat Commun. 2021. PMID: 34230464 Free PMC article.
Cited by
-
Olfactory Communication via Microbiota: What Is Known in Birds?Genes (Basel). 2018 Jul 31;9(8):387. doi: 10.3390/genes9080387. Genes (Basel). 2018. PMID: 30065222 Free PMC article. Review.
-
An individual and a sex odor signature in kittiwakes?: study of the semiochemical composition of preen secretion and preen down feathers.Naturwissenschaften. 2011 Jul;98(7):615-24. doi: 10.1007/s00114-011-0809-9. Epub 2011 Jun 8. Naturwissenschaften. 2011. PMID: 21656004
-
Wax Ester Composition of Songbird Preen Oil Varies Seasonally and Differs between Sexes, Ages, and Populations.J Chem Ecol. 2019 Jan;45(1):37-45. doi: 10.1007/s10886-018-1033-2. Epub 2018 Nov 20. J Chem Ecol. 2019. PMID: 30456558
-
Pheromones and signature mixtures: defining species-wide signals and variable cues for identity in both invertebrates and vertebrates.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2010 Oct;196(10):685-700. doi: 10.1007/s00359-010-0564-y. Epub 2010 Aug 3. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2010. PMID: 20680632 Review.
-
Metabolic Profiling Reveals That the Olfactory Cues in the Duck Uropygial Gland Potentially Act as Sex Pheromones.Animals (Basel). 2022 Feb 9;12(4):413. doi: 10.3390/ani12040413. Animals (Basel). 2022. PMID: 35203121 Free PMC article.
References
-
- Amo L, Galvan I, Tomas G, Sanz JJ. Predator odour recognition and avoidance in a songbird. Functional Ecology. 2008;22:289–293.
-
- Ball GF, Balthazart J. Neuroendocrine mechanisms regulating reproductive cycles and reproductive behavior in birds. In: Pfaff DW, Arnold AP, Etgen AM, Fahrbach SE, Rubin RT, editors. Hormones, Brain and Behavior. Academic Press; San Diego: 2002. pp. 649–798.
-
- Ball GF, Tlemçani O, Balthazart J. Induction of the Zenk protein after sexual interactions in male Japanese quail. Neuroreport. 1997;8:2965–2970. - PubMed
-
- Balthazart J, Absil P. Identification of catecholaminergic inputs to and outputs from aromatase-containing brain areas of the Japanese quail by tract tracing combined with tyrosine hydroxylase immunocytochemistry. J Comp Neurol. 1997;382:401–428. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
