

Review
doi: 10.1016/j.pbiomolbio.2010.05.001.
Epub 2010 May 20.
Structure and function of SWI/SNF chromatin remodeling complexes and mechanistic implications for transcription
Affiliations
- PMID: 20493208
- PMCID: PMC2924208
- DOI: 10.1016/j.pbiomolbio.2010.05.001
Item in Clipboard
Review
Structure and function of SWI/SNF chromatin remodeling complexes and mechanistic implications for transcription
Prog Biophys Mol Biol.
2010 Jun-Jul.
Abstract
ATP-dependent chromatin remodeling complexes are specialized protein machinery able to restructure the nucleosome to make its DNA accessible during transcription, replication and DNA repair. During the past few years structural biologists have defined the architecture and dynamics of some of these complexes using electron microscopy, shedding light on the mechanisms of action of these important assemblies. In this paper we review the existing structural information on the SWI/SNF family of the ATP-dependent chromatin remodeling complexes, and discuss their mechanistic implications.
Published by Elsevier Ltd.
Figures
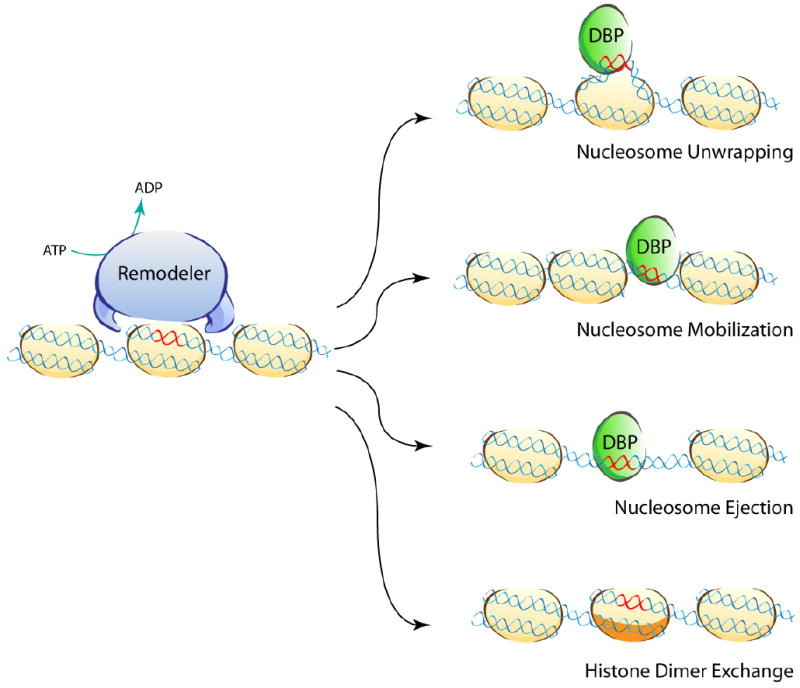
Different effects of the ATP dependent chromatin remodeling activity of remodelers on nucleosomal DNA: upon hydrolysis of ATP, a protected region of chromatin can become available to DNA binding protein complexes, such as transcription factors (in green). Nucleosomes can be unwrapped, mobilized or ejected to allow these processes. In some cases ATP dependent remodeling complexes can use ATP to introduce histone variants within the nucleosome by a process called dimer exchange. (DBP stands for DNA-binding protein)
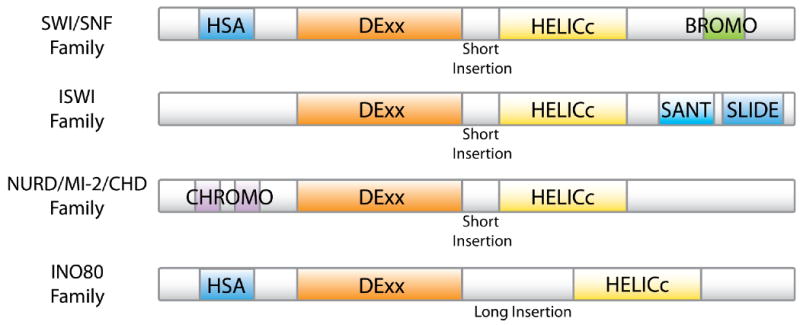
Classification of ATP-dependent chromatin remodeling complexes: The ATPase subunit of all the remodeling complexes belongs to the superfamily II helicase group. The ATPase always contains a DExx and a HELICc domain, spaced by a linker. The remodelers are classified into different families based on the presence of additional domains on their ATPase subunits. The SWI/SNF family contains a HSA domain, involved in actin binding, and a bromodomain important for the binding of acetylated lysines. The ISWI family contains the SANT and SLIDE domains, important for histone binding. The CHD/NURD/Mi-2 family contains a tandem Chromo domain, also used for histone binding. The INO80 family, like the SWI/SNF family, comprises a HSA domain but it is also characterized by the presence of a longer insertion between the DExx and the HELICc domains.
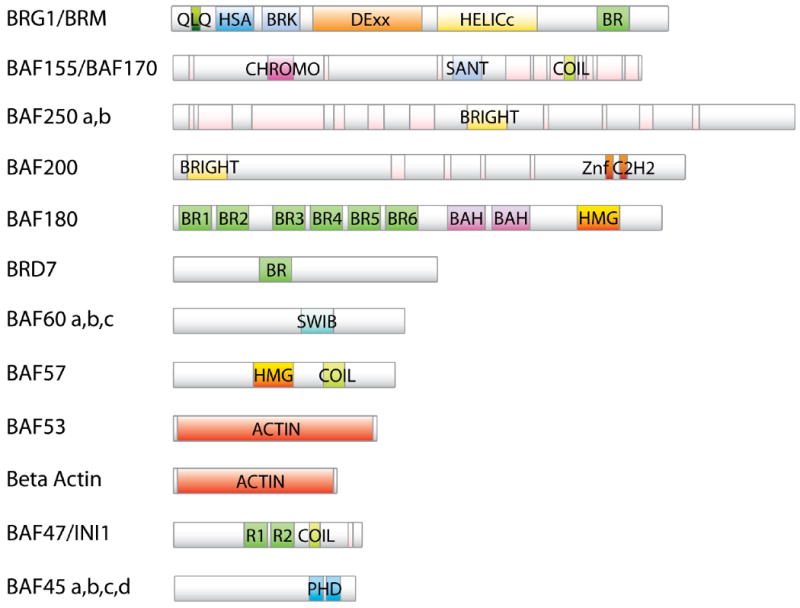
Prediction of the domain architecture of the human BAF and PBAF components. Proteins and domains are approximately in scale. Unlabeled pink modules indicate low complexity regions.
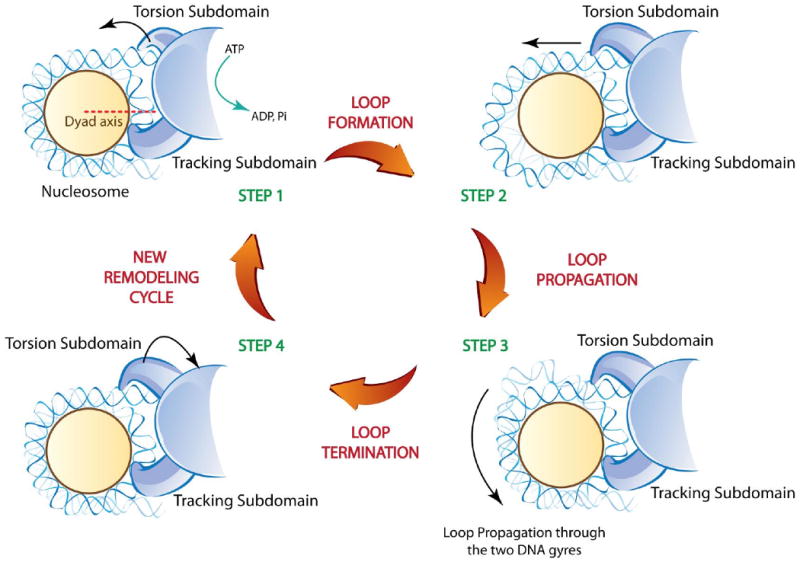
Schematic representation of the SWI/SNF dependent nucleosome remodeling process. The nucleosome is illustrated as spherical disc surrounded by the two DNA gyres (indicated with two different brightness based on the relative distance to the reader). Steps 1-4 indicate the different stages proposed to occur during the remodeling process. During Step1, the translocase domain binds the nucleosome two turns away from the dyad. Upon ATP-dependent hydrolysis, the torsion sub-domain generates a DNA loop that translocates through the tracking subdomain and the dyad, continuing in the second gyre (Step 2-3). The loop resolves when it reaches the exit site on the other side of the nucleosome (Step 4). The combination of these steps results in nucleosome repositioning. The complex is then ready for a new remodeling cycle (Step1).
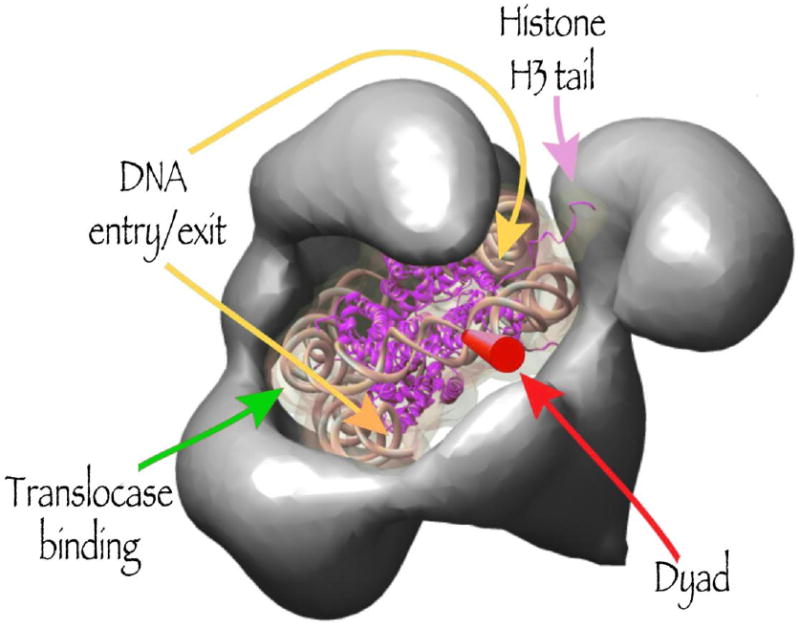
Functional model of nucleosome binding shown on the OTR reconstruction of the yeast RSC complex. Redrawn from Leschziner et al., 2007.
References
-
- Biedenkapp H, Borgmeyer U, Sippel AE, Klempnauer KH. Viral myb oncogene encodes a sequence-specific DNA-binding activity. Nature. 1998;335:835–7. - PubMed
-
- Boeger H, Griesenbeck J, Strattan JS, Kornberg RD. Nucleosomes unfold completely at a transcriptionally active promoter. Mol Cell. 2003;11:1587–98. - PubMed
-
- Cairns BR, Lorch Y, Li Y, Zhang M, Lacomis L, Erdjument-Bromage H, Tempst P, Du J, Laurent B, Kornberg RD. RSC, an essential, abundant chromatin-remodeling complex. Cell. 1996;87:1249–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
