

Coding in the mammalian gustatory system
- PMID: 20493563
- PMCID: PMC2902637
- DOI: 10.1016/j.tins.2010.04.002
Coding in the mammalian gustatory system
Abstract
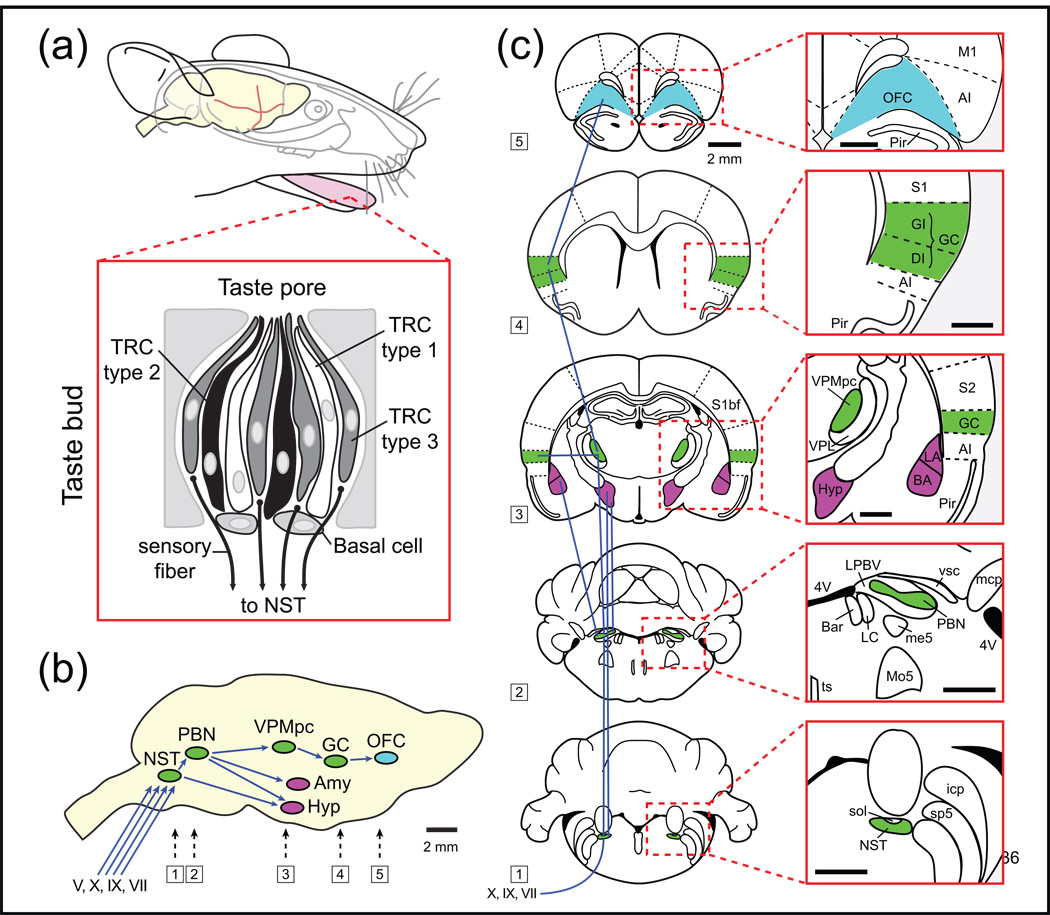
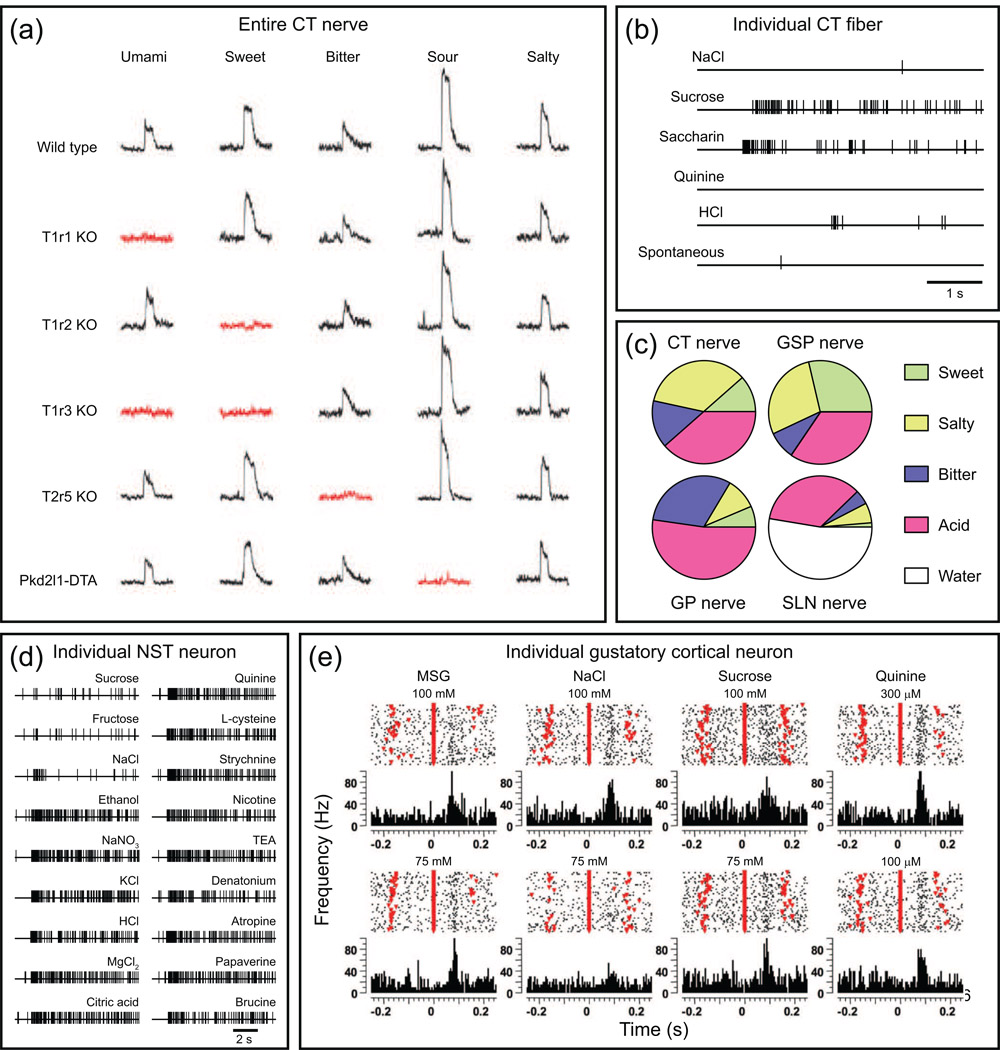
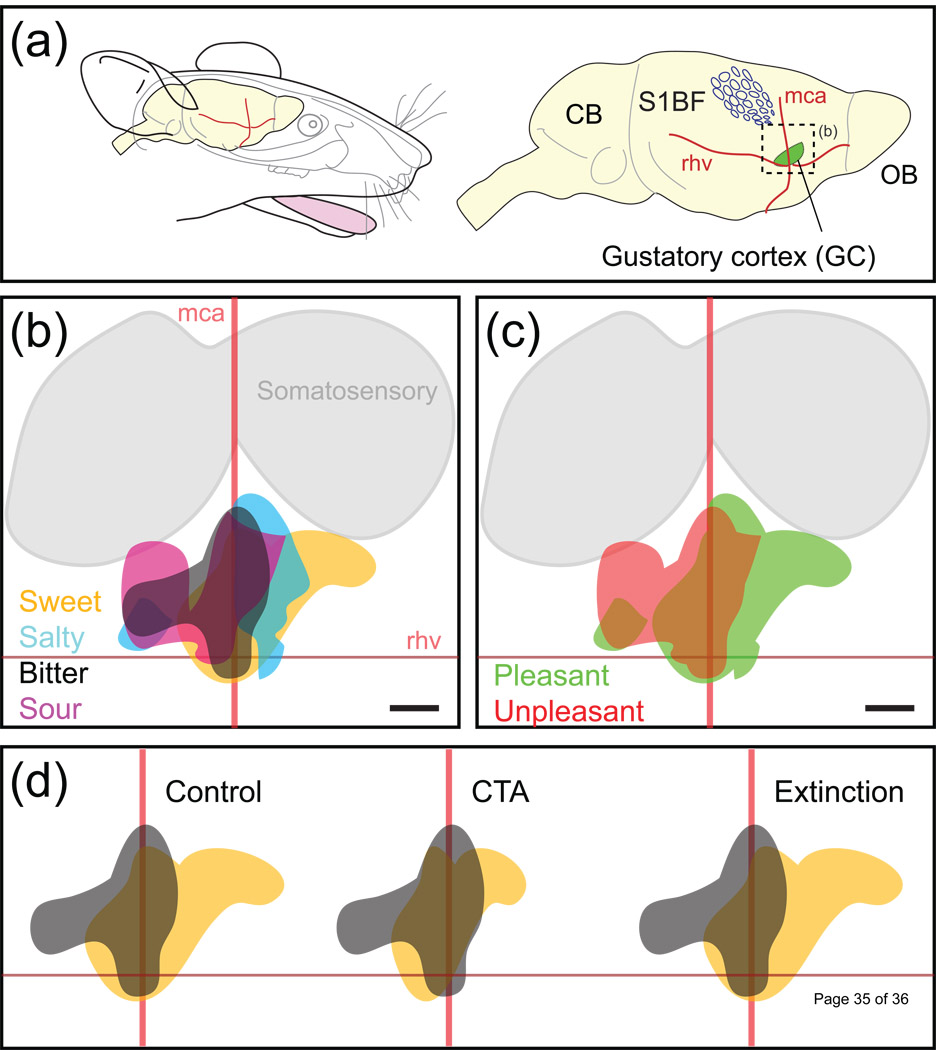
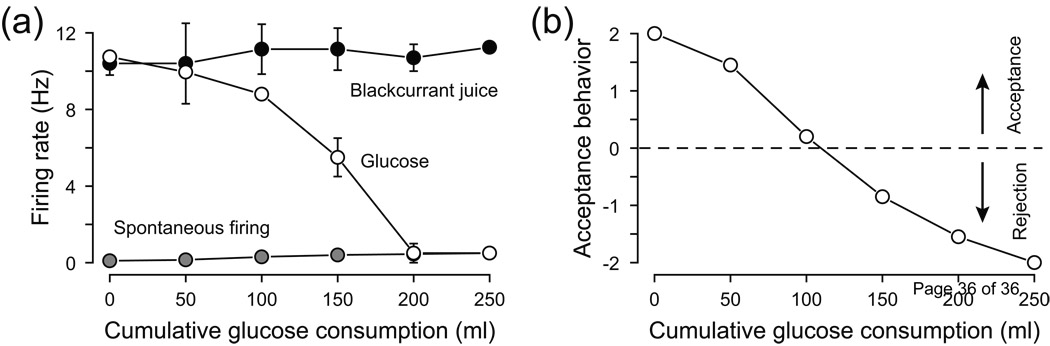
To understand gustatory physiology and associated dysfunctions it is important to know how oral taste stimuli are encoded both in the periphery and in taste-related brain centres. The identification of distinct taste receptors, together with electrophysiological recordings and behavioral assessments in response to taste stimuli, suggest that information about distinct taste modalities (e.g. sweet versus bitter) are transmitted from the periphery to the brain via segregated pathways. By contrast, gustatory neurons throughout the brain are more broadly tuned, indicating that ensembles of neurons encode taste qualities. Recent evidence reviewed here suggests that the coding of gustatory stimuli is not immutable, but is dependant on a variety of factors including appetite-regulating molecules and associative learning.
Copyright (c) 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Dalton P, et al. The merging of the senses: integration of subthreshold taste and smell. Nature neuroscience. 2000;3:431–432. - PubMed
-
- Gottfried JA, Dolan RJ. The nose smells what the eye sees: crossmodal visual facilitation of human olfactory perception. Neuron. 2003;39:375–386. - PubMed
-
- Spector AC, Travers SP. The representation of taste quality in the mammalian nervous system. Behav Cogn Neurosci Rev. 2005;4:143–191. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
