

S6K1 plays a critical role in early adipocyte differentiation
- PMID: 20493810
- PMCID: PMC2918254
- DOI: 10.1016/j.devcel.2010.02.018
S6K1 plays a critical role in early adipocyte differentiation
Abstract
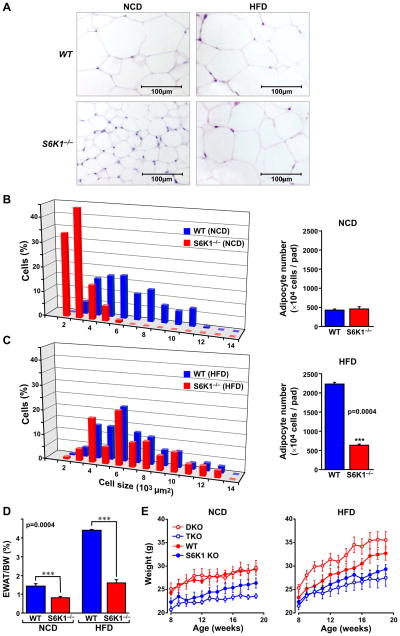
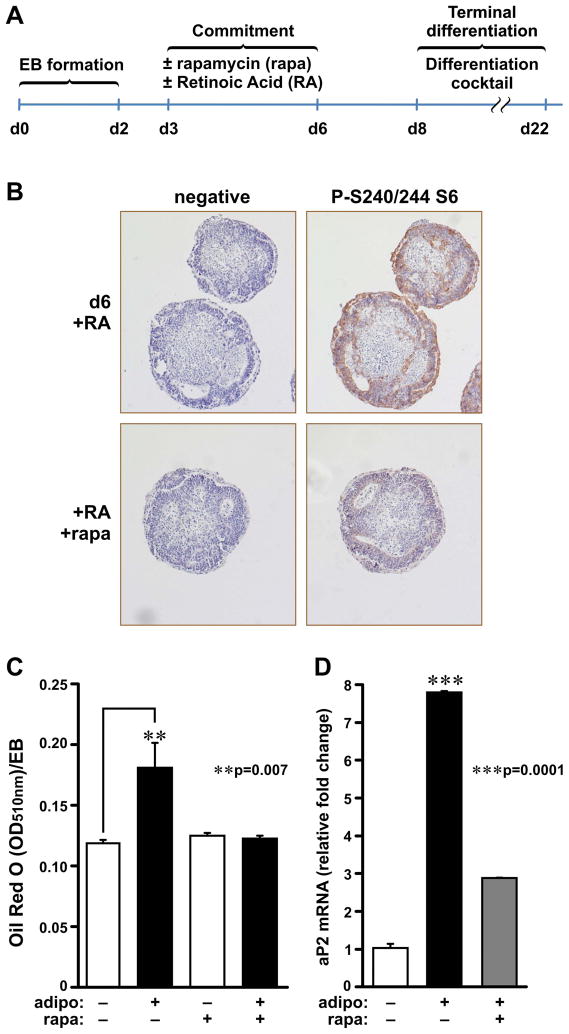
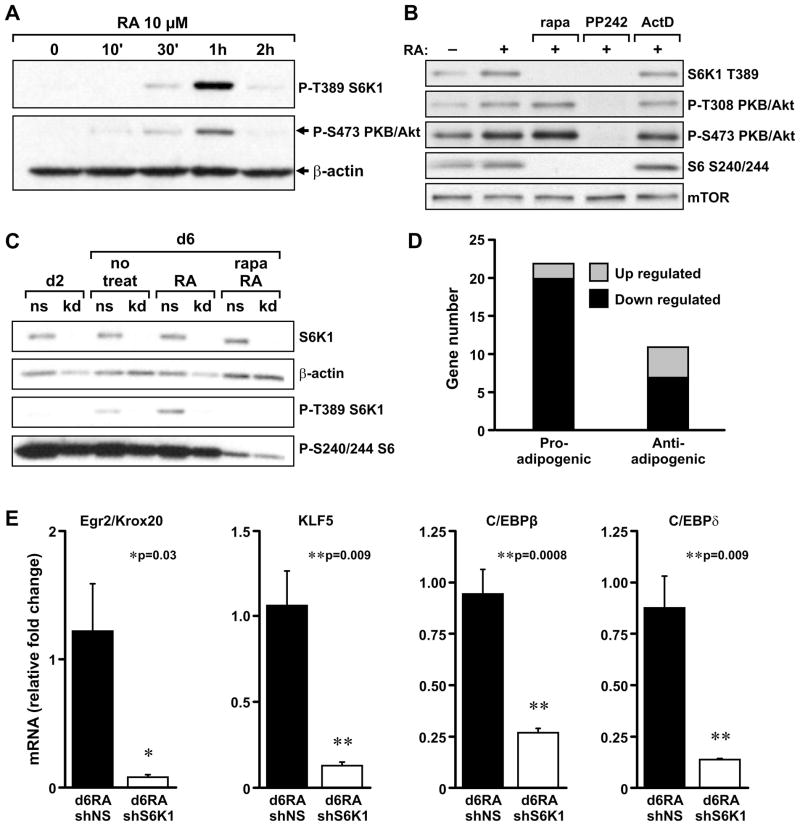
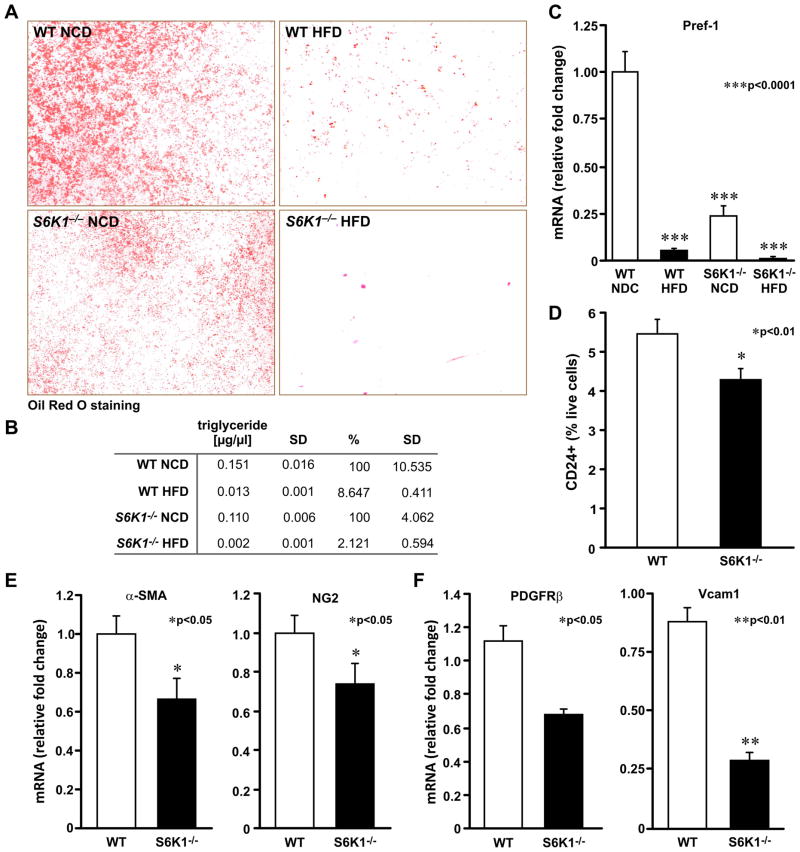
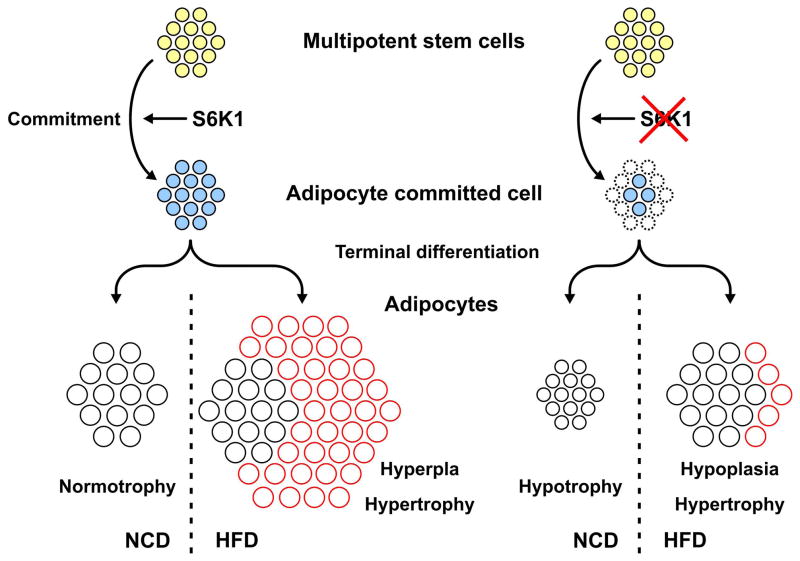
Earlier, we reported that S6K1(-/-) mice have reduced body fat mass, have elevated rates of lipolysis, have severely decreased adipocyte size, and are resistant to high fat diet (HFD)-induced obesity. Here we report that adipocytes of S6K1(-/-) mice on a HFD have the capacity to increase in size to a degree comparable to that of wild-type (WT) mice, but not in number, indicating an unexpected lesion in adipogenesis. Tracing this lesion revealed that S6K1 is dispensable for terminal adipocyte differentiation, but is involved in the commitment of embryonic stem cells to early adipocyte progenitors. We further show that absence of S6K1 attenuates the upregulation of transcription factors critical for commitment to adipogenesis. These results led to the conclusion that a lack of S6K1 impairs the generation of de novo adipocytes when mice are challenged with a HFD, consistent with a reduction in early adipocyte progenitors.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures


References
-
- Aguilar V, Alliouachene S, Sotiropoulos A, Sobering A, Athea Y, Djouadi F, Miraux S, Thiaudiere E, Foretz M, Viollet B, et al. S6 Kinase Deletion Suppresses Muscle Growth Adaptations to Nutrient Availability by Activating AMP Kinase. Cell Metab. 2007;5:476–487. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
