

Neural substrates underlying vestibular compensation: contribution of peripheral versus central processing
- PMID: 20495234
- PMCID: PMC3319765
- DOI: 10.3233/VES-2009-0357
Neural substrates underlying vestibular compensation: contribution of peripheral versus central processing
Abstract
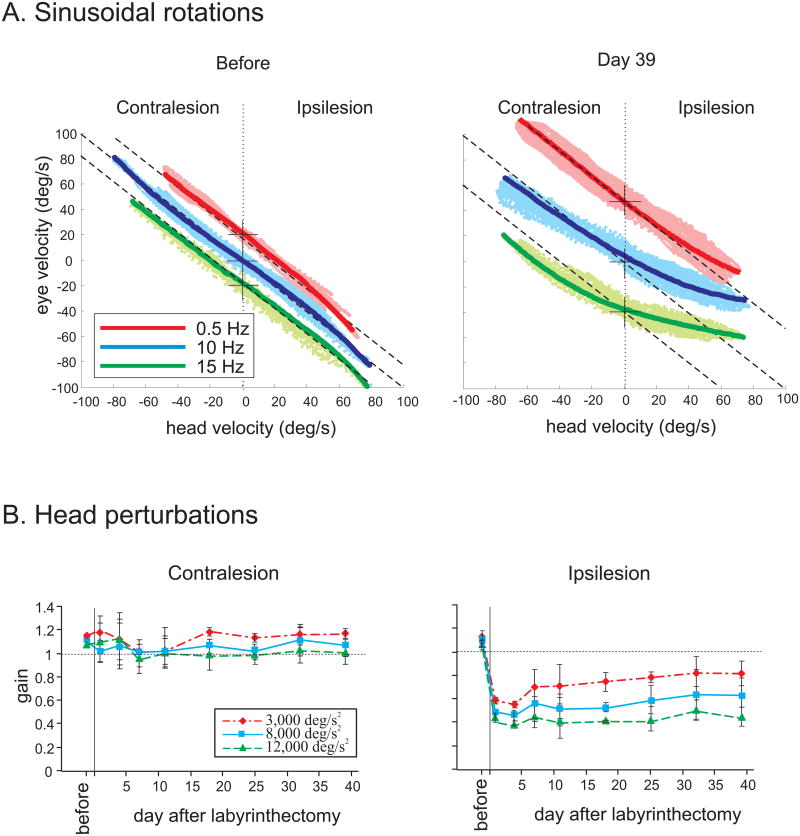
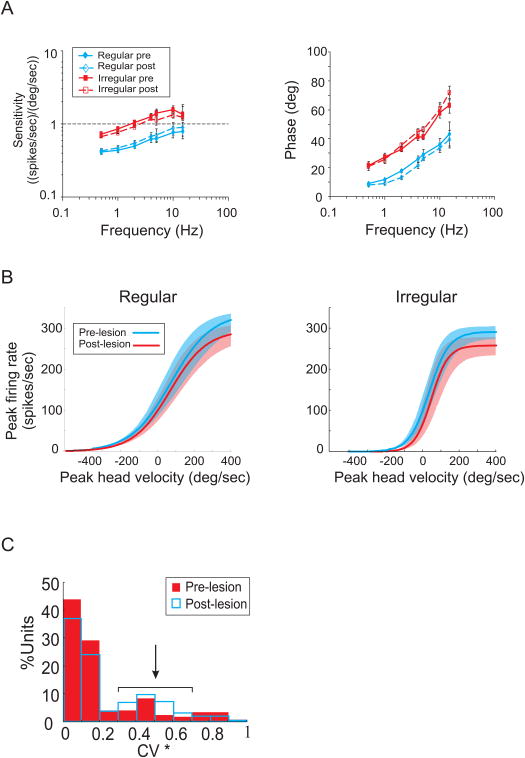
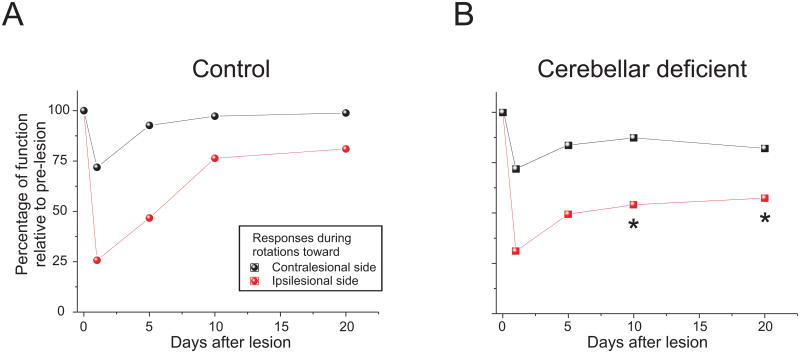
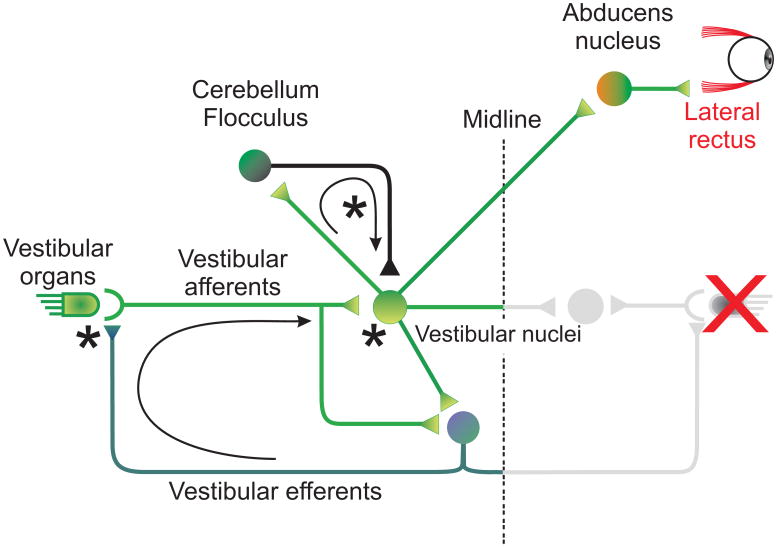
The vestibulo-ocular reflex (VOR), which functions to stabilize gaze and ensure clear vision during everyday activities, shows impressive adaptation in response to environmental requirements. In particular, the VOR exhibits remarkable recovery following the loss of unilateral labyrinthine input as a result of injury or disease. The relative simplicity of the pathways that mediate the VOR, make it an excellent model system for understanding the changes (learning) that occur in the brain following peripheral vestibular loss to yield adaptive changes. This mini review considers the findings of behavioral, single unit recording and lesion studies of VOR compensation. Recent experiments have provided evidence that the brain makes use of multiple plasticity mechanisms (i.e., changes in peripheral as well as central processing) during the course of vestibular compensation to accomplish the sensory-motor transformations required to accurately guide behavior.
Figures
References
-
- Aleisa M, Zeitouni AG, Cullen KE. Vestibular compensation after unilateral labyrinthectomy: normal versus cerebellar dysfunctional mice. J Otolaryngol. 2007;36:315–321. - PubMed
-
- Allum JH, Yamane M, Pfaltz CR. Long-term modifications of vertical and horizontal vestibuloocular reflex dynamics in man. I. After acute unilateral peripheral vestibular paralysis. Acta Otolaryngol. 1988;105:328–337. - PubMed
-
- Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodal sense. Annu Rev Neurosci. 2008;31:125–150. - PubMed
-
- Armand M, Minor LB. Relationship between time- and frequency-domain analyses of angular head movements in the squirrel monkey. J Comput Neurosci. 2001;11:217–239. - PubMed
-
- Baird RA, Desmadryl G, Fernandez C, Goldberg JM. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J Neurophysiol. 1988;60:182–203. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
