

Review
doi: 10.1038/nsmb.1843.
Epub 2010 May 23.
Signal integration through blending, bolstering and bifurcating of intracellular information
Affiliations
- PMID: 20495562
- PMCID: PMC3086636
- DOI: 10.1038/nsmb.1843
Item in Clipboard
Review
Signal integration through blending, bolstering and bifurcating of intracellular information
Nat Struct Mol Biol.
2010 Jun.
Abstract
A cell's response to its environment is often determined by signaling through the actions of enzyme cascades. The ability to organize these enzymes into multiprotein complexes allows for a high degree of fidelity, efficiency and spatial precision in signaling responses.
Figures
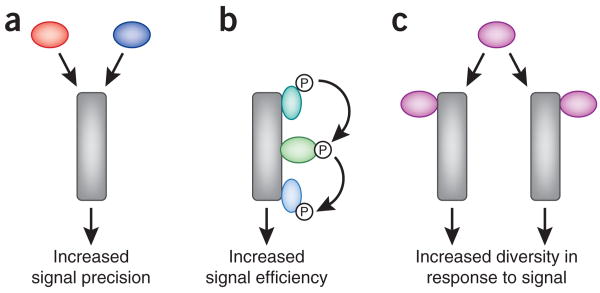
Schematic representation of mechanisms of signal transduction. (a) In signal integration, two independent inputs act on the same pathway to elicit a common outcome. (b) Scaffolding proteins allow the efficient relay of signals through successive enzymes in a pathway. (c) Specificity of signaling through broad-specificity enzymes is often assured by the spatial segregation of enzyme complexes.
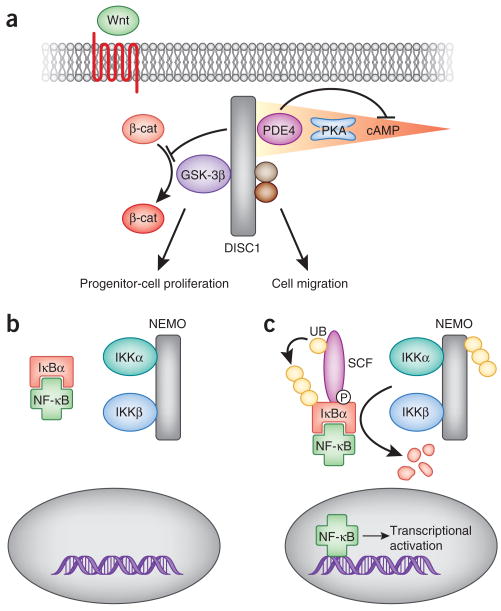
Signal integration: the DISC1 scaffold integrates Wnt and cAMP signaling to coordinate neurodevelopment. (a) DISC1 is required to regulate GSK-3β activity downstream of Wnt signaling, resulting in the stabilization and nuclear translocation of β-catenin (β-cat). This action of DISC1 is required for the regulation of progenitor-cell proliferation in the brain. Additionally, DISC1 coordinates PKA targets nuclear distribution protein nudE–like 1 (NDEL1) and LIS1 as well as PDE4 on the same molecular scaffold. This arrangement ensures tight control of cAMP-mediated signaling and likely has a role in cell migration. (b) Signaling through NF-κB requires the integration of phosphorylation and ubiquitination events. Under basal conditions, the nuclear localization signal (NLS) of NF-κB is masked by the binding of IκBs, holding NF-κB in the cytosol. (c) The nuclear translocation of NF-κB is initiated by the phosphorylation and ubiquitination of IκBs as a result of NEMO ubiquitination, IKK activation and recruitment of SCF E3 ligases to pIκB. UB, ubiquitin.
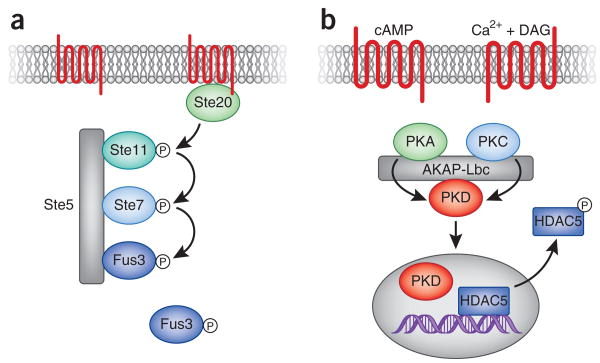
Ste5 coordinates signal activation in the yeast mating pathway. (a) In response to the presence of pheromone, Ste20 is activated at the cell membrane and in turn activates its downstream target Ste11. This initiates a cascade of phosphorylation events leading to the activation and release of the terminal component Fus3 from the Ste5 scaffold. By spatially coordinating the kinases involved in the pheromone response, Ste5 ensures both the speed and accuracy of signaling. (b) AKAP-Lbc coordinates the activation and translocation of protein kinase D. The integration of calcium/phospholipid (diacylglycerol, DAG) and cAMP signals promotes the activation of PKA and PKC respectively. These enzyme activities act synergistically to promote the phosphorylation of PKD in the active-site loop and its release from the anchoring protein complex. The newly active enzyme is then free to translocate into the nucleus, where it can phosphorylate the DNA-modifying enzyme HDAC5. Phospho-HDAC5 then exits the nucleus, thereby promoting the relaxation of DNA. In cardiomyocytes, this leads to transcriptional activation of genes that propagate the hypertrophic responses.
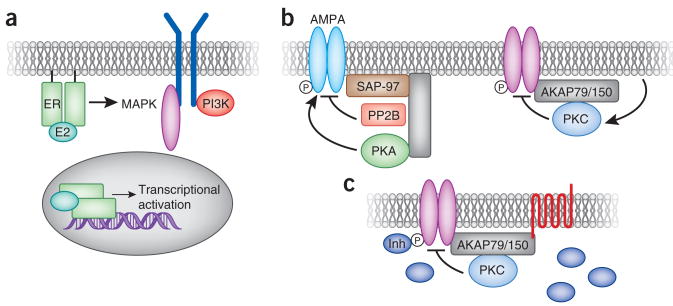
Segregation of estrogen signaling events. (a) Estrogen binds receptors at the plasma membrane and in the nucleus, leading to distinct cellular outcomes. Interaction with plasma-membrane receptors leads to cross-activation of other signaling components, including growth factor receptors, MAPK and PI 3-kinase (PI3K). Interaction with nuclear receptors leads to transcriptional activation of estrogen-responsive genes. Segregation of AKAP79/150 signaling events. (b) AKAP79/150 is cross-linked to the AMPA-type glutamate receptors via interaction with the membrane associated guanylate kinase (MAGUK) scaffolding protein SAP-97. This configuration brings the kinase PKA and the phosphatase PP2B in proximity to the channel to control its phosphorylation state. A complex between the same anchoring protein and a different binding partner, PKC, facilitates the phosphorylation-dependent suppression of M channels. (c) An unusual property of the AKAP79/150–PKC interaction is that the anchored kinase is rendered insensitive to ATP-competitive inhibitors (Inh). This creates a local pool of the enzyme that is resistant to these pharmacological agents.
References
-
- Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414:799–806. - PubMed
-
- Bers DM. Calcium cycling and signaling in cardiac myocytes. Annu Rev Physiol. 2008;70:23–49. - PubMed
-
- Millar JK, et al. Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum Mol Genet. 2000;9:1415–1423. - PubMed
-
- St Clair D, et al. Association within a family of a balanced autosomal translocation with major mental illness. Lancet. 1990;336:13–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
