

Structural basis of instability of the nucleosome containing a testis-specific histone variant, human H3T
- PMID: 20498094
- PMCID: PMC2890842
- DOI: 10.1073/pnas.1003064107
Structural basis of instability of the nucleosome containing a testis-specific histone variant, human H3T
Abstract
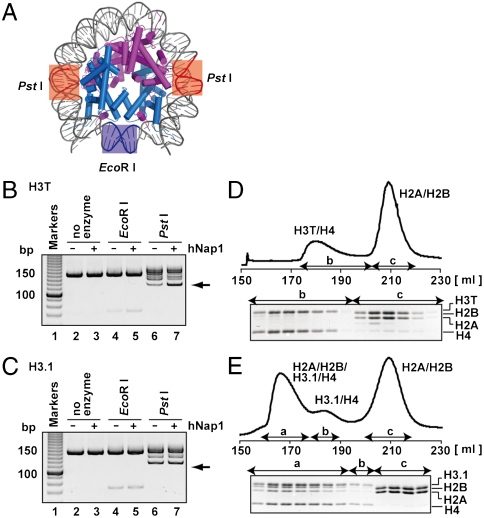
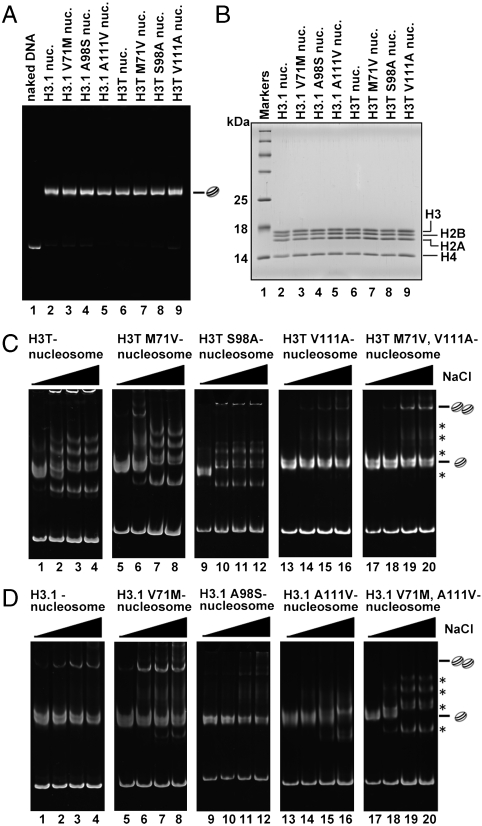
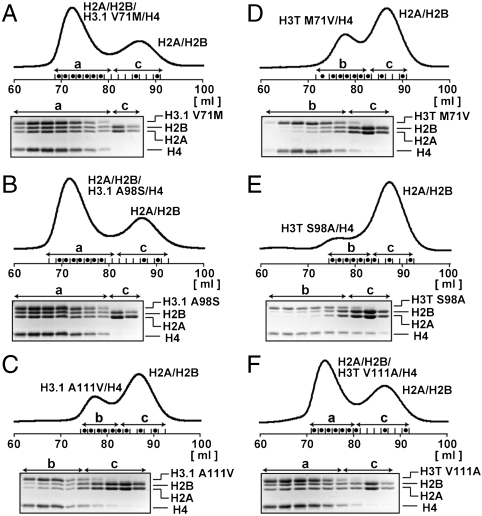
A histone H3 variant, H3T, is highly expressed in the testis, suggesting that it may play an important role in the chromatin reorganization required for meiosis and/or spermatogenesis. In the present study, we found that the nucleosome containing human H3T is significantly unstable both in vitro and in vivo, as compared to the conventional nucleosome containing H3.1. The crystal structure of the H3T nucleosome revealed structural differences in the H3T regions on both ends of the central alpha2 helix, as compared to those of H3.1. The H3T-specific residues (Met71 and Val111) are the source of the structural differences observed between H3T and H3.1. A mutational analysis revealed that these residues are responsible for the reduced stability of the H3T-containing nucleosome. These physical and structural properties of the H3T-containing nucleosome may provide the basis of chromatin reorganization during spermatogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Govin J, Caron C, Lestrat C, Rousseaux S, Khochbin S. The role of histones in chromatin remodelling during mammalian spermiogenesis. Eur J Biochem. 2004;271:3459–3469. - PubMed
-
- Franklin SG, Zweidler A. Non-allelic variants of histones 2a, 2b and 3 in mammals. Nature. 1977;266:273–275. - PubMed
-
- Albig W, Ebentheuer J, Klobeck G, Kunz J, Doenecke D. A solitary human H3 histone gene on chromosome 1. Hum Genet. 1996;97:486–491. - PubMed
-
- Witt O, Albig W, Doenecke D. Testis-specific expression of a novel human H3 histone gene. Exp Cell Res. 1996;229:301–306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
