

Interplay between DNA methylation and transcription factor availability: implications for developmental activation of the mouse Myogenin gene
- PMID: 20498275
- PMCID: PMC2916394
- DOI: 10.1128/MCB.00050-10
Interplay between DNA methylation and transcription factor availability: implications for developmental activation of the mouse Myogenin gene
Abstract
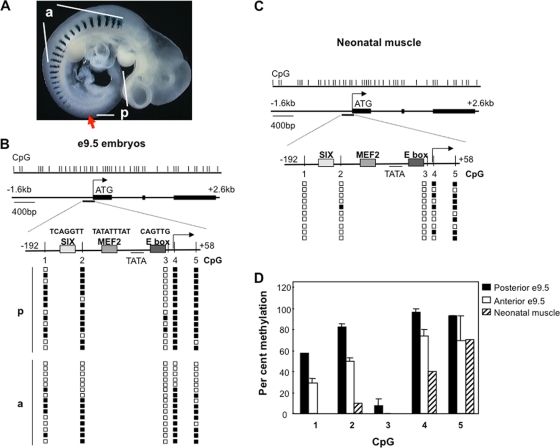
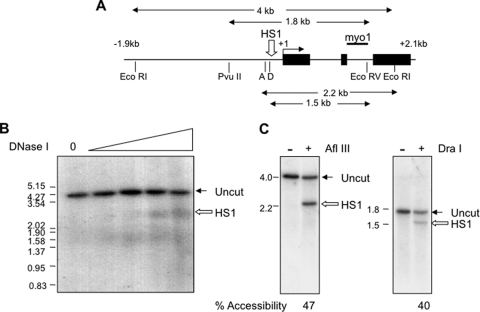
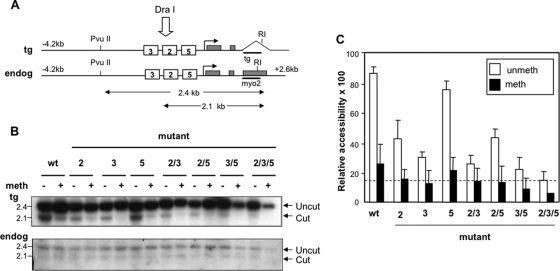
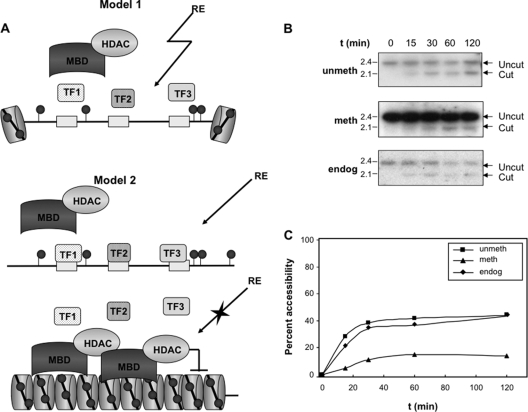
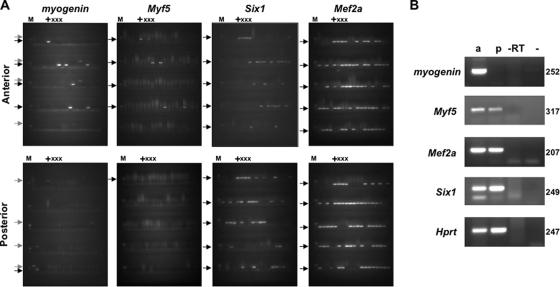
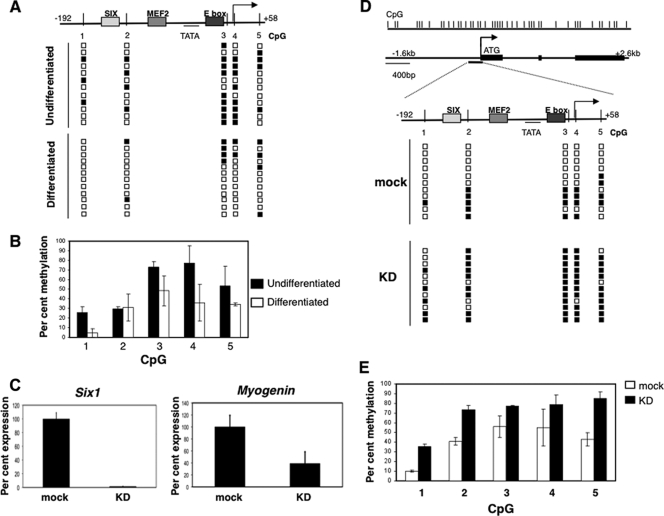
During development, gene activation is stringently regulated to restrict expression only to the correct cell type and correct developmental stage. Here, we present mechanistic evidence that suggests DNA methylation contributes to this regulation by suppressing premature gene activation. Using the mouse Myogenin promoter as an example of the weak CpG island class of promoters, we find that it is initially methylated but becomes demethylated as development proceeds. Full hypersensitive site formation of the Myogenin promoter requires both the MEF2 and SIX binding sites, but binding to only one site can trigger the partial chromatin opening of the nonmethylated promoter. DNA methylation markedly decreases hypersensitive site formation that now occurs at a detectable level only when binding to both MEF2 and SIX binding sites is possible. This suggests that the probability of activating the methylated promoter is low until two of the factors are coexpressed within the same cell. Consistent with this, the single-cell analysis of developing somites shows that the coexpression of MEF2A and SIX1, which bind the MEF2 and SIX sites, correlates with the fraction of cells that demethylate the Myogenin promoter. Taken together, these studies imply that DNA methylation helps to prevent inappropriate gene activation until sufficient activating factors are coexpressed.
Figures
Similar articles
-
Myogenin and MEF2 function synergistically to activate the MRF4 promoter during myogenesis.Mol Cell Biol. 1995 May;15(5):2707-18. doi: 10.1128/MCB.15.5.2707. Mol Cell Biol. 1995. PMID: 7739551 Free PMC article.
-
Transcriptional activation of the myogenin gene by MEF2-mediated recruitment of myf5 is inhibited by adenovirus E1A protein.Biochem Biophys Res Commun. 1999 Nov;265(1):222-32. doi: 10.1006/bbrc.1999.1390. Biochem Biophys Res Commun. 1999. PMID: 10548518
-
Myocyte enhancer factor 2C and myogenin up-regulate each other's expression and induce the development of skeletal muscle in P19 cells.J Biol Chem. 2000 Jan 7;275(1):41-6. doi: 10.1074/jbc.275.1.41. J Biol Chem. 2000. PMID: 10617583
-
Synergistic up-regulation of muscle LIM protein expression in C2C12 and NIH3T3 cells by myogenin and MEF2C.Mol Genet Genomics. 2009 Jan;281(1):1-10. doi: 10.1007/s00438-008-0393-7. Epub 2008 Nov 6. Mol Genet Genomics. 2009. PMID: 18987887 Review.
-
Combinatorial control of muscle development by basic helix-loop-helix and MADS-box transcription factors.Proc Natl Acad Sci U S A. 1996 Sep 3;93(18):9366-73. doi: 10.1073/pnas.93.18.9366. Proc Natl Acad Sci U S A. 1996. PMID: 8790335 Free PMC article. Review.
Cited by
-
Mitochondrial Function in Muscle Stem Cell Fates.Front Cell Dev Biol. 2020 Jun 16;8:480. doi: 10.3389/fcell.2020.00480. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32612995 Free PMC article. Review.
-
Turning on myogenin in muscle: a paradigm for understanding mechanisms of tissue-specific gene expression.Comp Funct Genomics. 2012;2012:836374. doi: 10.1155/2012/836374. Epub 2012 Jun 28. Comp Funct Genomics. 2012. PMID: 22811619 Free PMC article.
-
DNA Methylation Dynamics During Differentiation, Proliferation, and Tumorigenesis in the Intestinal Tract.Stem Cells Dev. 2015 Dec 1;24(23):2733-9. doi: 10.1089/scd.2015.0235. Epub 2015 Oct 20. Stem Cells Dev. 2015. PMID: 26413818 Free PMC article. Review.
-
An Origanum majorana Leaf Diet Influences Myogenin Gene Expression, Performance, and Carcass Characteristics in Lambs.Animals (Basel). 2022 Dec 20;13(1):14. doi: 10.3390/ani13010014. Animals (Basel). 2022. PMID: 36611623 Free PMC article.
-
Global DNA methylation pattern involved in the modulation of differentiation potential of adipogenic and myogenic precursors in skeletal muscle of pigs.Stem Cell Res Ther. 2020 Dec 11;11(1):536. doi: 10.1186/s13287-020-02053-3. Stem Cell Res Ther. 2020. PMID: 33308295 Free PMC article.
References
-
- Antequera, F., J. Boyes, and A. Bird. 1990. High levels of de novo methylation and altered chromatin structure at CpG islands in cell lines. Cell 62:503-514. - PubMed
-
- Berkes, C. A., D. A. Bergstrom, B. H. Penn, K. J. Seaver, P. S. Knoepfler, and S. J. Tapscott. 2004. Pbx marks genes for activation by MyoD indicating a role for a homeodomain protein in establishing myogenic potential. Mol. Cell 14:465-477. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases