

Neisseria meningitidis Opc invasin binds to the sulphated tyrosines of activated vitronectin to attach to and invade human brain endothelial cells
- PMID: 20502634
- PMCID: PMC2873925
- DOI: 10.1371/journal.ppat.1000911
Neisseria meningitidis Opc invasin binds to the sulphated tyrosines of activated vitronectin to attach to and invade human brain endothelial cells
Abstract
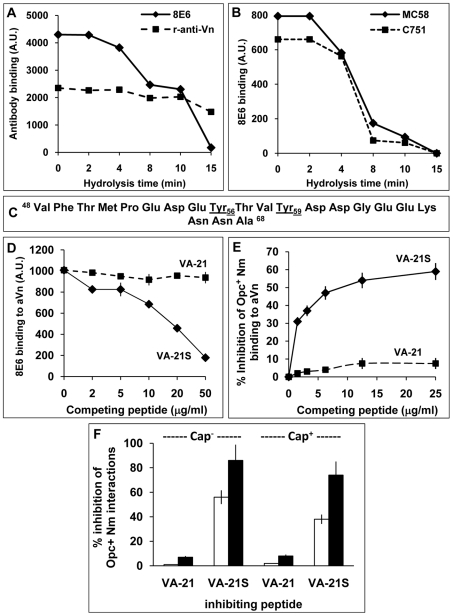
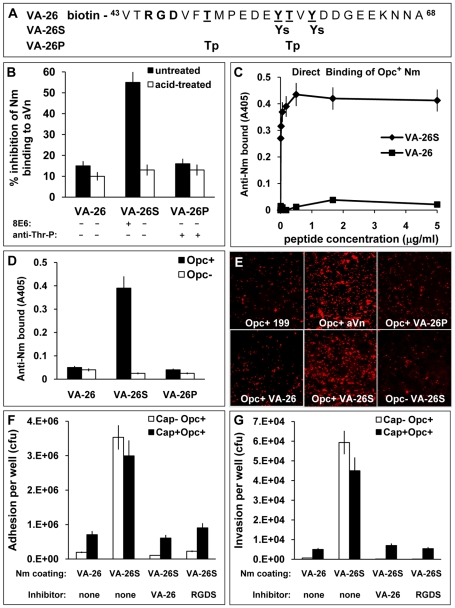
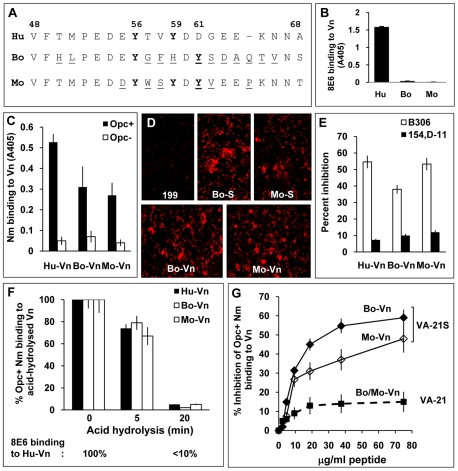
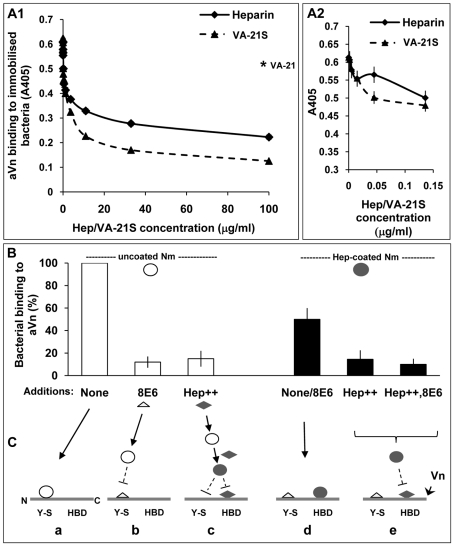
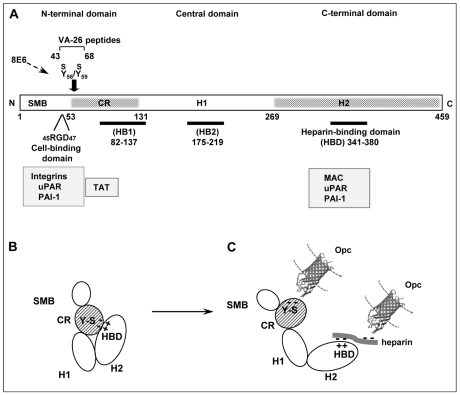
The host vasculature is believed to constitute the principal route of dissemination of Neisseria meningitidis (Nm) throughout the body, resulting in septicaemia and meningitis in susceptible humans. In vitro, the Nm outer membrane protein Opc can enhance cellular entry and exit, utilising serum factors to anchor to endothelial integrins; but the mechanisms of binding to serum factors are poorly characterised. This study demonstrates that Nm Opc expressed in acapsulate as well as capsulate bacteria can increase human brain endothelial cell line (HBMEC) adhesion and entry by first binding to serum vitronectin and, to a lesser extent, fibronectin. This study also demonstrates that Opc binds preferentially to the activated form of human vitronectin, but not to native vitronectin unless the latter is treated to relax its closed conformation. The direct binding of vitronectin occurs at its Connecting Region (CR) requiring sulphated tyrosines Y(56) and Y(59). Accordingly, Opc/vitronectin interaction could be inhibited with a conformation-dependent monoclonal antibody 8E6 that targets the sulphotyrosines, and with synthetic sulphated (but not phosphorylated or unmodified) peptides spanning the vitronectin residues 43-68. Most importantly, the 26-mer sulphated peptide bearing the cell-binding domain (45)RGD(47) was sufficient for efficient meningococcal invasion of HBMECs. To our knowledge, this is the first study describing the binding of a bacterial adhesin to sulphated tyrosines of the host receptor. Our data also show that a single region of Opc is likely to interact with the sulphated regions of both vitronectin and of heparin. As such, in the absence of heparin, Opc-expressing Nm interact directly at the CR but when precoated with heparin, they bind via heparin to the heparin-binding domain of the activated vitronectin, although with a lower affinity than at the CR. Such redundancy suggests the importance of Opc/vitronectin interaction in meningococcal pathogenesis and may enable the bacterium to harness the benefits of the physiological processes in which the host effector molecule participates.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Visualisation and quantification of intracellular interactions of Neisseria meningitidis and human α-actinin by confocal imaging.J Vis Exp. 2010 Oct 24;(44):2045. doi: 10.3791/2045. J Vis Exp. 2010. PMID: 21085092 Free PMC article.
-
Interaction of Neisseria meningitidis with human brain microvascular endothelial cells: role of MAP- and tyrosine kinases in invasion and inflammatory cytokine release.Cell Microbiol. 2004 Dec;6(12):1153-66. doi: 10.1111/j.1462-5822.2004.00422.x. Cell Microbiol. 2004. PMID: 15527495
-
Distinct mechanisms of interactions of Opc-expressing meningococci at apical and basolateral surfaces of human endothelial cells; the role of integrins in apical interactions.Mol Microbiol. 1994 Oct;14(1):173-84. doi: 10.1111/j.1365-2958.1994.tb01277.x. Mol Microbiol. 1994. PMID: 7530316
-
Molecular mechanisms involved in the interaction of Neisseria meningitidis with cells of the human blood-cerebrospinal fluid barrier.Pathog Dis. 2017 Mar 1;75(2). doi: 10.1093/femspd/ftx023. Pathog Dis. 2017. PMID: 28334198 Review.
-
Interactions of meningococcal virulence factors with endothelial cells at the human blood-cerebrospinal fluid barrier and their role in pathogenicity.FEBS Lett. 2016 Nov;590(21):3854-3867. doi: 10.1002/1873-3468.12344. Epub 2016 Aug 23. FEBS Lett. 2016. PMID: 27498906 Review.
Cited by
-
Two strikingly different signaling pathways are induced by meningococcal type IV pili on endothelial and epithelial cells.Infect Immun. 2012 Jan;80(1):175-86. doi: 10.1128/IAI.05837-11. Epub 2011 Nov 7. Infect Immun. 2012. PMID: 22064711 Free PMC article.
-
Small Rho GTPases and the Effector VipA Mediate the Invasion of Epithelial Cells by Filamentous Legionella pneumophila.Front Cell Infect Microbiol. 2018 May 3;8:133. doi: 10.3389/fcimb.2018.00133. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 29774203 Free PMC article.
-
Unraveling Neisseria meningitidis pathogenesis: from functional genomics to experimental models.F1000Res. 2017 Jul 26;6:1228. doi: 10.12688/f1000research.11279.1. eCollection 2017. F1000Res. 2017. PMID: 28781769 Free PMC article. Review.
-
Host-pathogen interactions in bacterial meningitis.Acta Neuropathol. 2016 Feb;131(2):185-209. doi: 10.1007/s00401-015-1531-z. Epub 2016 Jan 7. Acta Neuropathol. 2016. PMID: 26744349 Free PMC article. Review.
-
Polar invasion and translocation of Neisseria meningitidis and Streptococcus suis in a novel human model of the blood-cerebrospinal fluid barrier.PLoS One. 2012;7(1):e30069. doi: 10.1371/journal.pone.0030069. Epub 2012 Jan 11. PLoS One. 2012. PMID: 22253884 Free PMC article.
References
-
- Virji M, Makepeace K, Moxon ER. Distinct mechanisms of interactions of Opc-expressing meningococci at apical and basolateral surfaces of human endothelial cells; the role of integrins in apical interactions. Mol Microbiol. 1994;14:173–184. - PubMed
-
- Virji M, Makepeace K, Peak IR, Ferguson DJ, Jennings MP, et al. Opc- and pilus-dependent interactions of meningococci with human endothelial cells: molecular mechanisms and modulation by surface polysaccharides. Mol Microbiol. 1995;18:741–754. - PubMed
-
- Virji M, Makepeace K, Ferguson DJ, Achtman M, Sarkari J, et al. Expression of the Opc protein correlates with invasion of epithelial and endothelial cells by Neisseria meningitidis. Mol Microbiol. 1992;6:2785–2795. - PubMed
-
- Unkmeir A, Latsch K, Dietrich G, Wintermeyer E, Schinke B, et al. Fibronectin mediates Opc-dependent internalization of Neisseria meningitidis in human brain microvascular endothelial cells. Mol Microbiol. 2002;46:933–946. - PubMed
-
- Lambotin M, Hoffmann I, Laran-Chich MP, Nassif X, Couraud PO, et al. Invasion of endothelial cells by Neisseria meningitidis requires cortactin recruitment by a phosphoinositide-3-kinase/Rac1 signalling pathway triggered by the lipo-oligosaccharide. J Cell Sci. 2005;118:3805–3816. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
