

The Plasmodium falciparum-specific human memory B cell compartment expands gradually with repeated malaria infections
- PMID: 20502681
- PMCID: PMC2873912
- DOI: 10.1371/journal.ppat.1000912
The Plasmodium falciparum-specific human memory B cell compartment expands gradually with repeated malaria infections
Abstract
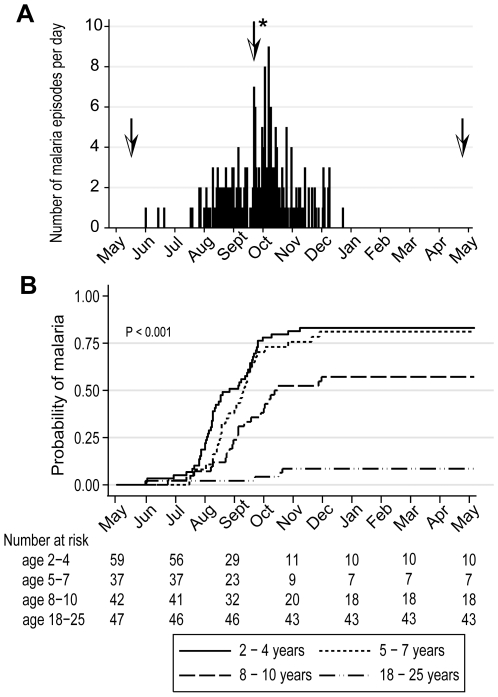
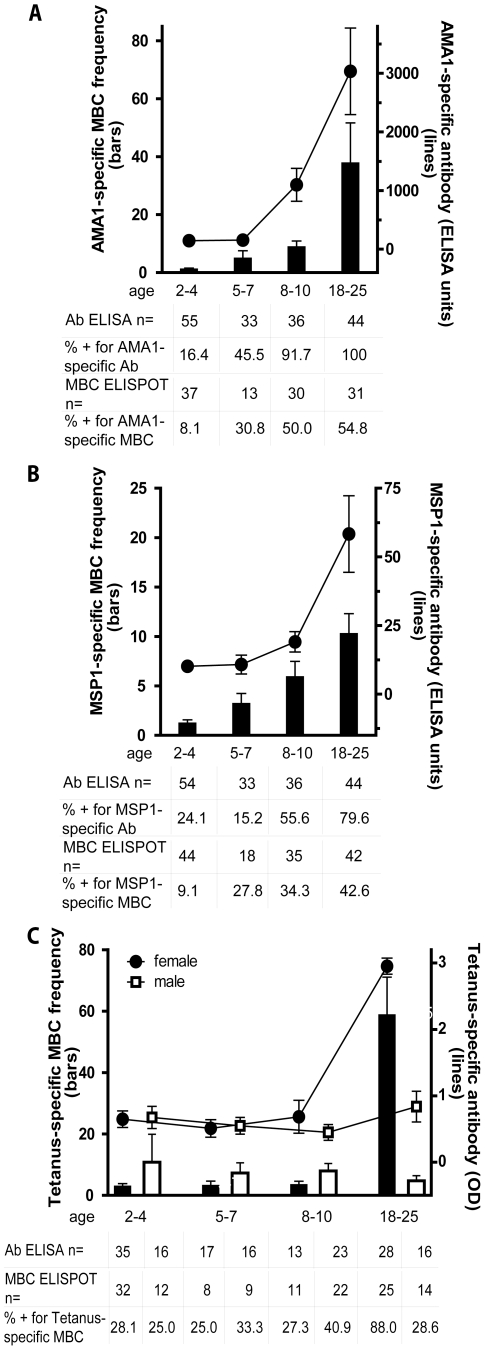
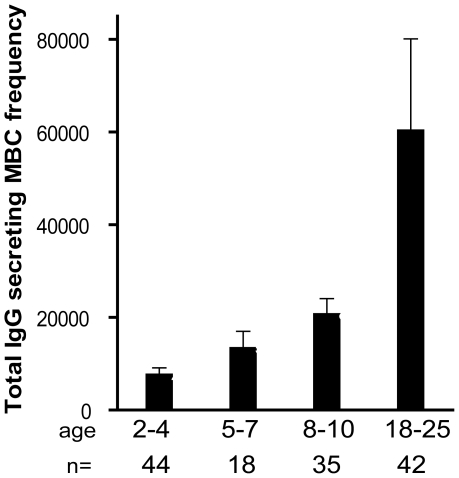
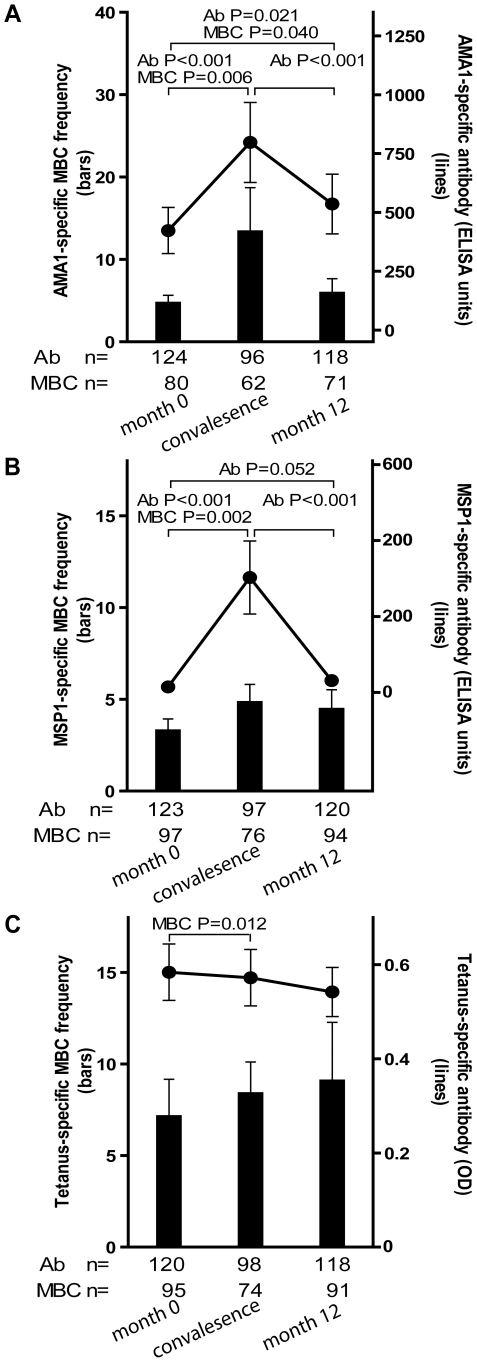
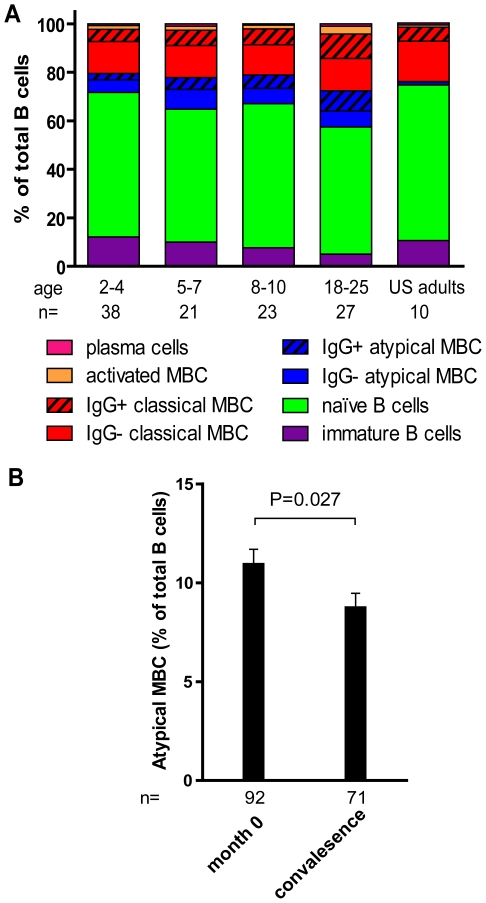
Immunity to Plasmodium falciparum (Pf) malaria is only acquired after years of repeated infections and wanes rapidly without ongoing parasite exposure. Antibodies are central to malaria immunity, yet little is known about the B-cell biology that underlies the inefficient acquisition of Pf-specific humoral immunity. This year-long prospective study in Mali of 185 individuals aged 2 to 25 years shows that Pf-specific memory B-cells and antibodies are acquired gradually in a stepwise fashion over years of repeated Pf exposure. Both Pf-specific memory B cells and antibody titers increased after acute malaria and then, after six months of decreased Pf exposure, contracted to a point slightly higher than pre-infection levels. This inefficient, stepwise expansion of both the Pf-specific memory B-cell and long-lived antibody compartments depends on Pf exposure rather than age, based on the comparator response to tetanus vaccination that was efficient and stable. These observations lend new insights into the cellular basis of the delayed acquisition of malaria immunity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Plotkin SA. Vaccines: correlates of vaccine-induced immunity. Clin Infect Dis. 2008;47:401–409. - PubMed
-
- Vekemans J, Ballou WR. Plasmodium falciparum malaria vaccines in development. Expert Rev Vaccines. 2008;7:223–240. - PubMed
-
- Marsh K, Kinyanjui S. Immune effector mechanisms in malaria. Parasite Immunol. 2006;28:51–60. - PubMed
-
- Cohen S, Mc GI, Carrington S. Gamma-globulin and acquired immunity to human malaria. Nature. 1961;192:733–737. - PubMed
-
- Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol. 2008;9:725–732. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
