

EBV promotes human CD8 NKT cell development
- PMID: 20502687
- PMCID: PMC2873918
- DOI: 10.1371/journal.ppat.1000915
EBV promotes human CD8 NKT cell development
Retraction in
-
Retraction: EBV promotes human CD8 NKT cell development.PLoS Pathog. 2010 Dec 3;6(12):10.1371/annotation/1730f52d-2ba9-4f08-b330-47d71b31ae4a. doi: 10.1371/annotation/1730f52d-2ba9-4f08-b330-47d71b31ae4a. PLoS Pathog. 2010. PMID: 21151944 Free PMC article. No abstract available.
Abstract
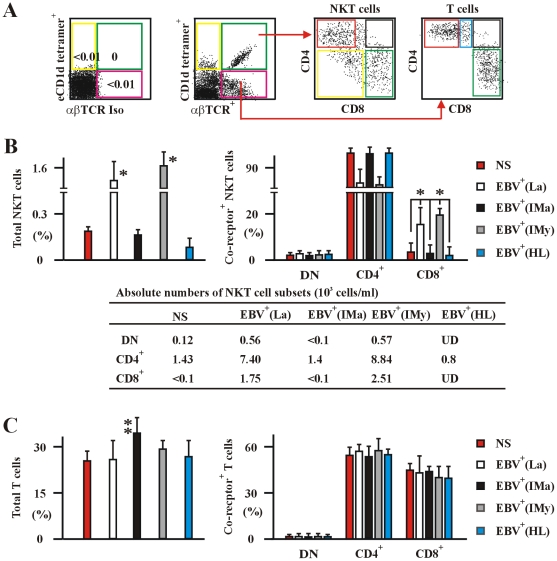
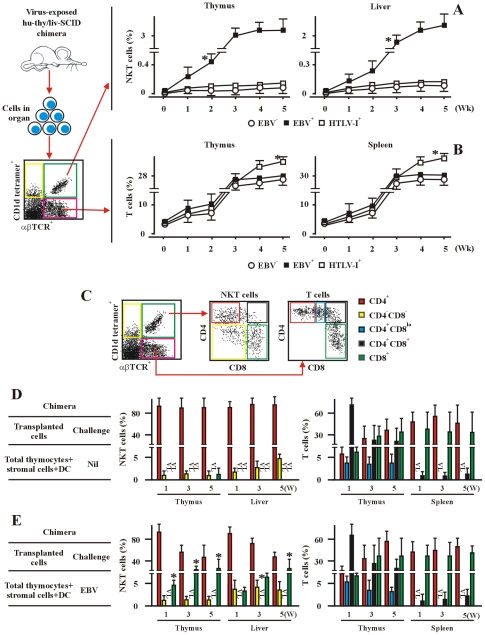
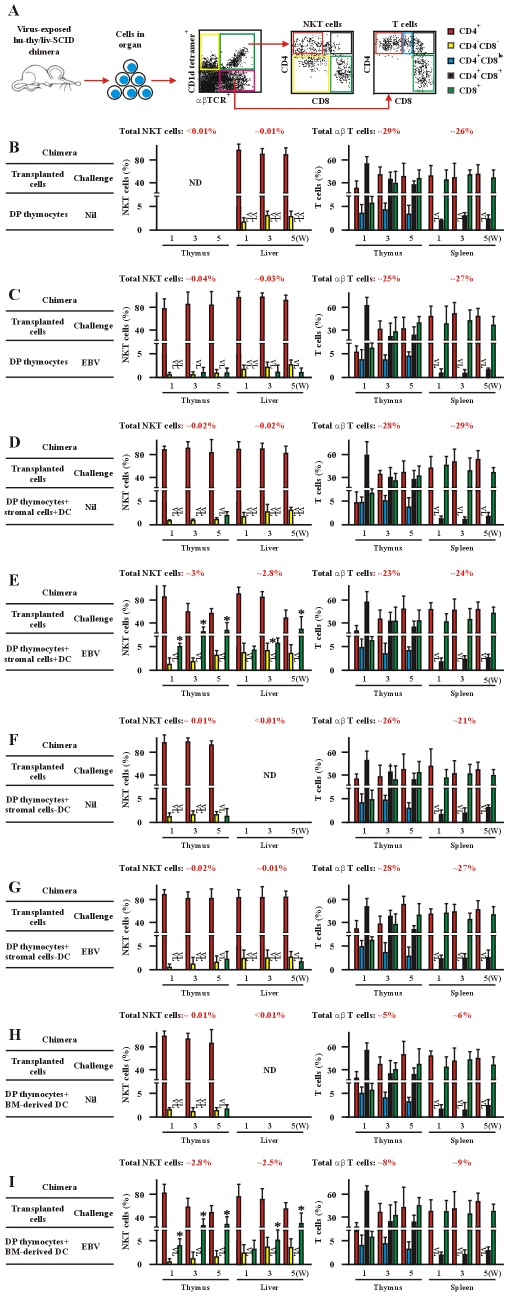
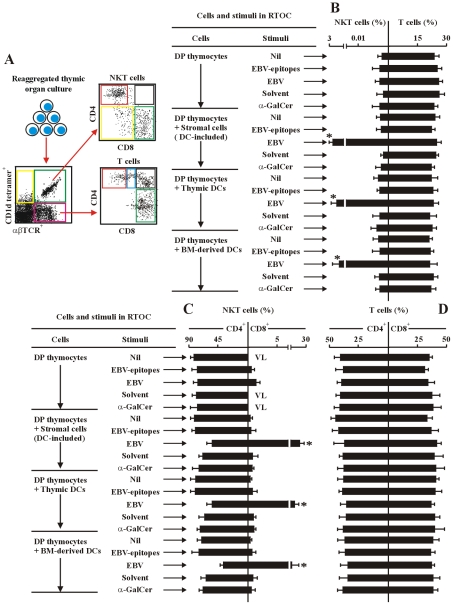
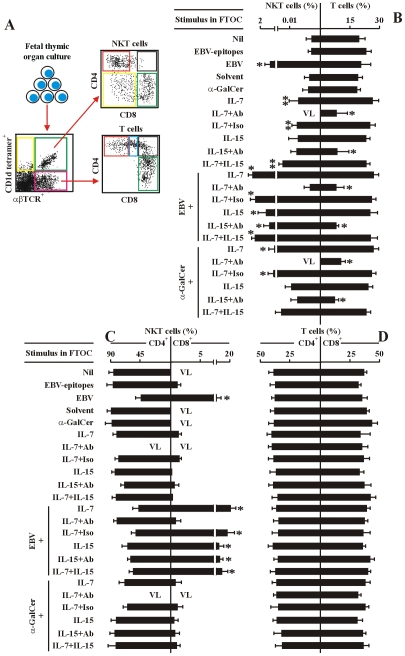
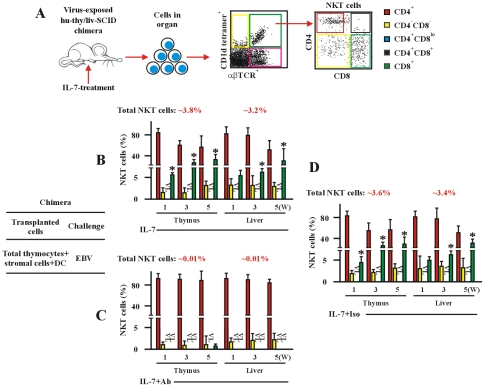
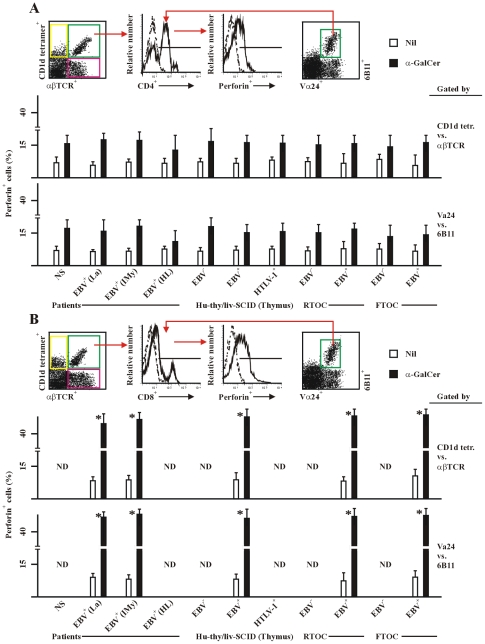
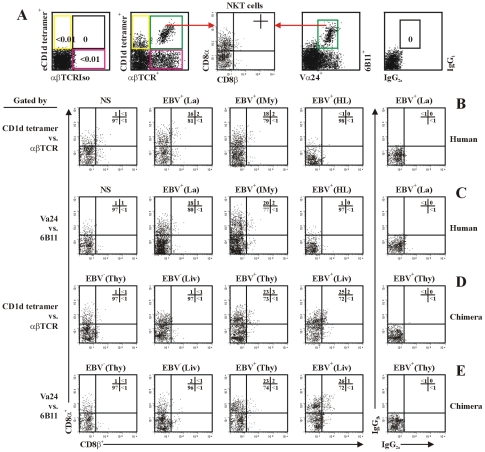
The reports on the origin of human CD8(+) Valpha24(+) T-cell receptor (TCR) natural killer T (NKT) cells are controversial. The underlying mechanism that controls human CD4 versus CD8 NKT cell development is not well-characterized. In the present study, we have studied total 177 eligible patients and subjects including 128 healthy latent Epstein-Barr-virus(EBV)-infected subjects, 17 newly-onset acute infectious mononucleosis patients, 16 newly-diagnosed EBV-associated Hodgkin lymphoma patients, and 16 EBV-negative normal control subjects. We have established human-thymus/liver-SCID chimera, reaggregated thymic organ culture, and fetal thymic organ culture. We here show that the average frequency of total and CD8(+) NKT cells in PBMCs from 128 healthy latent EBV-infected subjects is significantly higher than in 17 acute EBV infectious mononucleosis patients, 16 EBV-associated Hodgkin lymphoma patients, and 16 EBV-negative normal control subjects. However, the frequency of total and CD8(+) NKT cells is remarkably increased in the acute EBV infectious mononucleosis patients at year 1 post-onset. EBV-challenge promotes CD8(+) NKT cell development in the thymus of human-thymus/liver-SCID chimeras. The frequency of total (3% of thymic cells) and CD8(+) NKT cells ( approximately 25% of NKT cells) is significantly increased in EBV-challenged chimeras, compared to those in the unchallenged chimeras (<0.01% of thymic cells, CD8(+) NKT cells undetectable, respectively). The EBV-induced increase in thymic NKT cells is also reflected in the periphery, where there is an increase in total and CD8(+) NKT cells in liver and peripheral blood in EBV-challenged chimeras. EBV-induced thymic CD8(+) NKT cells display an activated memory phenotype (CD69(+)CD45RO(hi)CD161(+)CD62L(lo)). After EBV-challenge, a proportion of NKT precursors diverges from DP thymocytes, develops and differentiates into mature CD8(+) NKT cells in thymus in EBV-challenged human-thymus/liver-SCID chimeras or reaggregated thymic organ cultures. Thymic antigen-presenting EBV-infected dendritic cells are required for this process. IL-7, produced mainly by thymic dendritic cells, is a major and essential factor for CD8(+) NKT cell differentiation in EBV-challenged human-thymus/liver-SCID chimeras and fetal thymic organ cultures. Additionally, these EBV-induced CD8(+) NKT cells produce remarkably more perforin than that in counterpart CD4(+) NKT cells, and predominately express CD8alphaalpha homodimer in their co-receptor. Thus, upon interaction with certain viruses, CD8 lineage-specific NKT cells are developed, differentiated and matured intrathymically, a finding with potential therapeutic importance against viral infections and tumors.
Conflict of interest statement
TJ, HY, XR, XW, LL, and WL have a patent pending on a method to establish the EBV-challenged human-thymus/liver-SCID chimera for NKT cell research and therapeutic usage.
Figures
Similar articles
-
EBV-induced human CD8+ NKT cells suppress tumorigenesis by EBV-associated malignancies.Cancer Res. 2009 Oct 15;69(20):7935-44. doi: 10.1158/0008-5472.CAN-09-0828. Epub 2009 Oct 6. Cancer Res. 2009. PMID: 19808969
-
EBV-induced human CD8(+) NKT cells synergise CD4(+) NKT cells suppressing EBV-associated tumours upon induction of Th1-bias.Cell Mol Immunol. 2009 Oct;6(5):367-79. doi: 10.1038/cmi.2009.48. Cell Mol Immunol. 2009. PMID: 19887050 Free PMC article.
-
Comparative immunophenotypic features of EBV-positive and EBV-negative atypical lymphocytosis.Cytometry B Clin Cytom. 2003 Sep;55(1):22-8. doi: 10.1002/cyto.b.10043. Cytometry B Clin Cytom. 2003. PMID: 12949956
-
Role of human natural killer cells during Epstein-Barr virus infection.Crit Rev Immunol. 2014;34(6):501-7. doi: 10.1615/critrevimmunol.2014012312. Crit Rev Immunol. 2014. PMID: 25597312 Review.
-
Thymic differentiation of TCR alpha beta(+) CD8 alpha alpha(+) IELs.Immunol Rev. 2007 Feb;215:178-88. doi: 10.1111/j.1600-065X.2006.00488.x. Immunol Rev. 2007. PMID: 17291288 Review.
Cited by
-
Comprehensive multi-omics single-cell data integration reveals greater heterogeneity in the human immune system.iScience. 2022 Sep 13;25(10):105123. doi: 10.1016/j.isci.2022.105123. eCollection 2022 Oct 21. iScience. 2022. PMID: 36185375 Free PMC article.
-
Epstein-barr virus, the immune system, and associated diseases.Front Microbiol. 2011 Jan 26;2:5. doi: 10.3389/fmicb.2011.00005. eCollection 2011. Front Microbiol. 2011. PMID: 21687403 Free PMC article.
-
Role of Innate T Cells in Anti-Bacterial Immunity.Front Immunol. 2015 Jun 11;6:302. doi: 10.3389/fimmu.2015.00302. eCollection 2015. Front Immunol. 2015. PMID: 26124758 Free PMC article. Review.
-
Exhaustion‑like dysfunction of T and NKT cells in an X‑linked severe combined immunodeficiency patient with maternal engraftment by single‑cell analysis.Int J Mol Med. 2023 Mar;51(3):25. doi: 10.3892/ijmm.2023.5228. Epub 2023 Feb 17. Int J Mol Med. 2023. PMID: 36799159 Free PMC article.
-
Testosterone increases susceptibility to amebic liver abscess in mice and mediates inhibition of IFNγ secretion in natural killer T cells.PLoS One. 2013;8(2):e55694. doi: 10.1371/journal.pone.0055694. Epub 2013 Feb 12. PLoS One. 2013. PMID: 23424637 Free PMC article.
References
-
- Kronenberg M. Toward an understanding of NKT cell biology: progress and paradoxes. Annu Rev Immunol. 2005;23:877–900. - PubMed
-
- Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297–336. - PubMed
-
- Brigl M, Brenner MB. CD1: antigen presentation and T cell function. Annu Rev Immunol. 2004;22:817–890. - PubMed
-
- Zhou D, Mattner J, Cantu C, 3rd, Schrantz N, Yin N, et al. Lysosomal glycosphingolipid recognition by NKT cells. Science. 2004;306:1786–1789. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
