

Mice lacking adrenergic signaling have normal cochlear responses and normal resistance to acoustic injury but enhanced susceptibility to middle-ear infection
- PMID: 20503062
- PMCID: PMC2914237
- DOI: 10.1007/s10162-010-0220-9
Mice lacking adrenergic signaling have normal cochlear responses and normal resistance to acoustic injury but enhanced susceptibility to middle-ear infection
Abstract
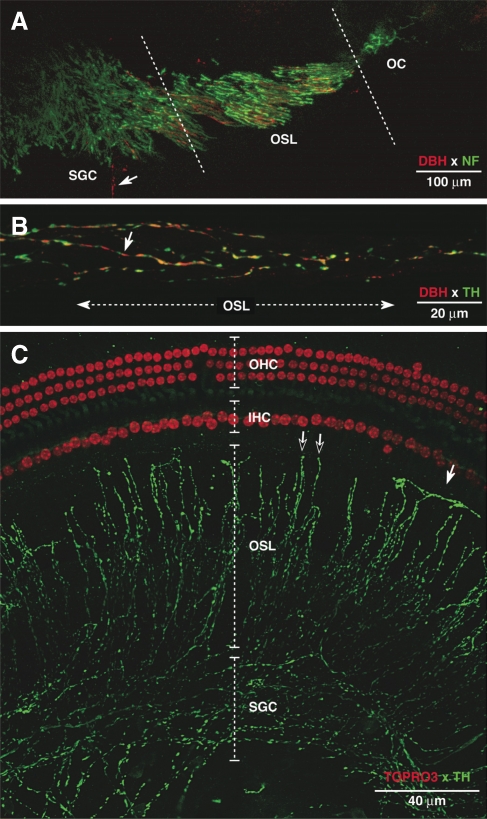
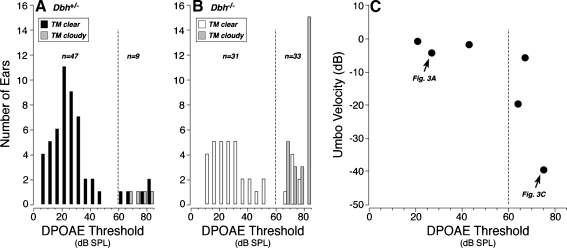
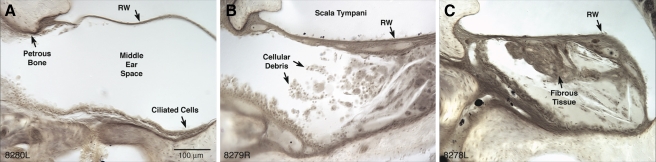
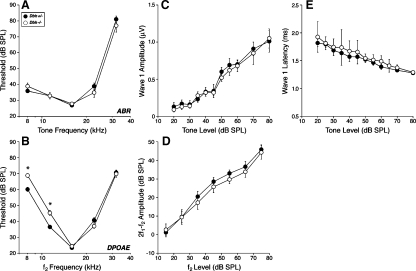
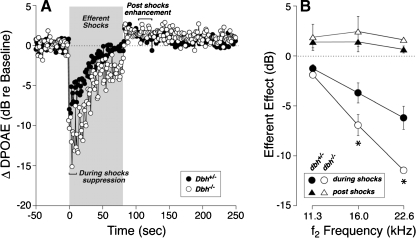
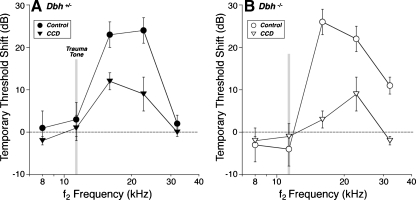
The vasculature and neurons of the inner ear receive adrenergic innervation from the cervical sympathetic chain, and adrenergic receptors may be expressed by cells of the organ of Corti and stria vascularis, despite a lack of direct sympathetic innervation. To assess the functional role of adrenergic signaling in the auditory periphery, we studied mice with targeted deletion of the gene for dopamine beta-hydroxylase (DBH), which catalyzes the conversion of dopamine to noradrenaline; thus, these mutant mice have no measurable adrenaline or noradrenaline. Dbh (-/-) mice were more susceptible to spontaneous middle-ear infection than their control littermates, consistent with a role for sympathetics in systemic and/or local immune response. At 6-8 weeks of age, cochlear thresholds and suprathreshold responses assessed by auditory brainstem responses and distortion product otoacoustic emissions, as well as light-microscopic morphology, were indistinguishable from controls, if ears with conductive hearing loss were eliminated. Dbh (-/-) mice were no more susceptible to acoustic injury than controls, despite prior reports that sympathectomy reduces noise damage. Dbh (-/-) mice showed enhancement of shock-evoked olivocochlear suppression of cochlear responses, which may arise from the loss of adrenergic inputs to olivocochlear neurons in the brainstem. However, adrenergic modulation of olivocochlear efferents does not mediate the protective effect of contralateral cochlear destruction on ipsilateral response to acoustic overexposure.
Figures


Similar articles
-
Muscarinic signaling in the cochlea: presynaptic and postsynaptic effects on efferent feedback and afferent excitability.J Neurosci. 2010 May 12;30(19):6751-62. doi: 10.1523/JNEUROSCI.5080-09.2010. J Neurosci. 2010. PMID: 20463237 Free PMC article.
-
Dopaminergic signaling in the cochlea: receptor expression patterns and deletion phenotypes.J Neurosci. 2012 Jan 4;32(1):344-55. doi: 10.1523/JNEUROSCI.4720-11.2012. J Neurosci. 2012. PMID: 22219295 Free PMC article.
-
Contralateral cochlear effects of ipsilateral damage: no evidence for interaural coupling.Hear Res. 2010 Feb;260(1-2):70-80. doi: 10.1016/j.heares.2009.11.011. Epub 2009 Nov 26. Hear Res. 2010. PMID: 19944141 Free PMC article.
-
Olivocochlear efferents: Their action, effects, measurement and uses, and the impact of the new conception of cochlear mechanical responses.Hear Res. 2018 May;362:38-47. doi: 10.1016/j.heares.2017.12.012. Epub 2017 Dec 21. Hear Res. 2018. PMID: 29291948 Free PMC article. Review.
-
The cochlea as an independent neuroendocrine organ: expression and possible roles of a local hypothalamic-pituitary-adrenal axis-equivalent signaling system.Hear Res. 2012 Jun;288(1-2):3-18. doi: 10.1016/j.heares.2012.03.007. Epub 2012 Mar 29. Hear Res. 2012. PMID: 22484018 Free PMC article. Review.
Cited by
-
Spontaneous Otoacoustic Emissions in TectaY1870C/+ Mice Reflect Changes in Cochlear Amplification and How It Is Controlled by the Tectorial Membrane.eNeuro. 2018 Dec 26;5(6):ENEURO.0314-18.2018. doi: 10.1523/ENEURO.0314-18.2018. eCollection 2018 Nov-Dec. eNeuro. 2018. PMID: 30627650 Free PMC article.
-
Understanding the aetiology and resolution of chronic otitis media from animal and human studies.Dis Model Mech. 2017 Nov 1;10(11):1289-1300. doi: 10.1242/dmm.029983. Dis Model Mech. 2017. PMID: 29125825 Free PMC article. Review.
-
Norepinephrine is necessary for experience-dependent plasticity in the developing mouse auditory cortex.J Neurosci. 2015 Feb 11;35(6):2432-7. doi: 10.1523/JNEUROSCI.0532-14.2015. J Neurosci. 2015. PMID: 25673838 Free PMC article.
-
Expression and localization of α2A-adrenergic receptor in the rat post-natal developing cochlea.Eur J Histochem. 2023 Aug 7;67(3):3748. doi: 10.4081/ejh.2023.3748. Eur J Histochem. 2023. PMID: 37548252 Free PMC article.
-
Norepinephrine protects against cochlear outer hair cell damage and noise-induced hearing loss via α2A-adrenergic receptor.BMC Neurosci. 2024 Jan 30;25(1):5. doi: 10.1186/s12868-024-00845-4. BMC Neurosci. 2024. PMID: 38291397 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
