

Neuropilin-1 interacts with the second branchial arch microenvironment to mediate chick neural crest cell dynamics
- PMID: 20503363
- PMCID: PMC3077279
- DOI: 10.1002/dvdy.22303
Neuropilin-1 interacts with the second branchial arch microenvironment to mediate chick neural crest cell dynamics
Abstract
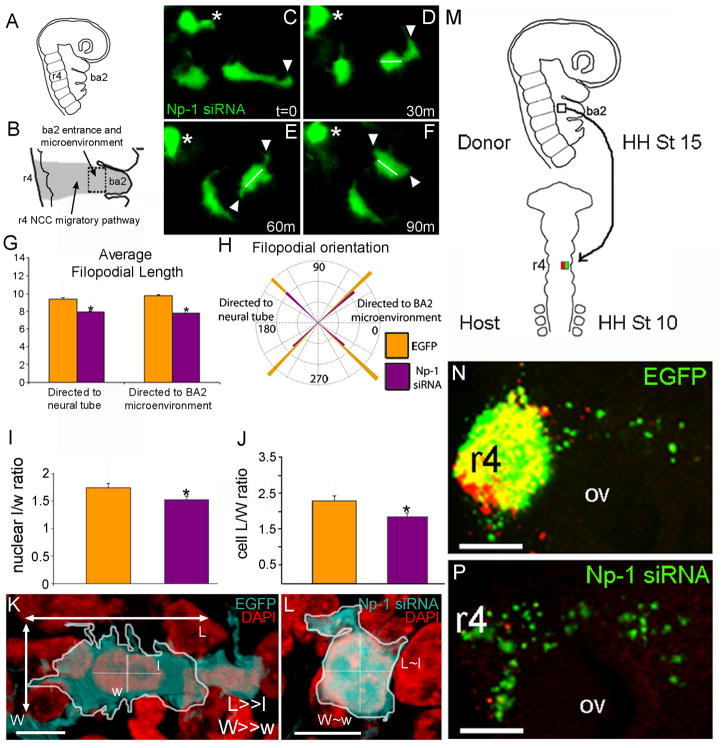
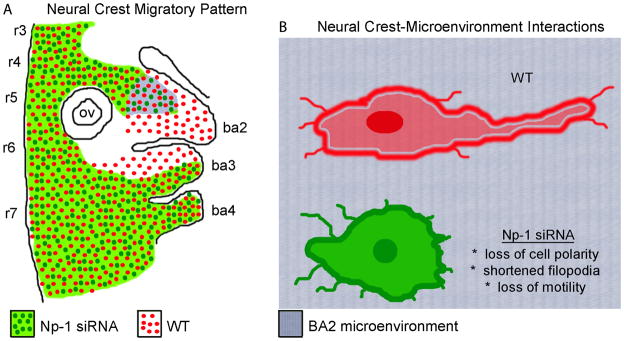
Cranial neural crest cells (NCCs) require neuropilin signaling to reach and invade the branchial arches. Here, we use an in vivo chick model to investigate whether the neuropilin-1 knockdown phenotype is specific to the second branchial arch (ba2), changes in NCC behaviors and phenotypic consequences, and whether neuropilins work together to facilitate entry into and invasion of ba2. We find that cranial NCCs with reduced neuropilin-1 expression displayed shorter protrusions and decreased cell body and nuclear length-to-width ratios characteristic of a loss in polarity and motility, after specific interaction with ba2. Directed NCC migration was rescued by transplantation of transfected NCCs into rhombomere 4 of younger hosts. Lastly, reduction of neuropilin-2 expression by shRNA either solely or with reduction of neuropilin-1 expression did not lead to a stronger head phenotype. Thus, NCCs, independent of rhombomere origin, require neuropilin-1, but not neuropilin-2 to maintain polarity and directed migration into ba2.
Figures



References
-
- Baker CV, Bronner-Fraser M. The origins of the neural crest. Part I: embryonic induction. Mech Dev. 1997;69:3–11. - PubMed
-
- Bron R, Eickholt BJ, Vermeren M, Fragale N, Cohen J. Functional knockdown of neuropilin-1 in the developing chick nervous system by siRNA hairpins phenocopies genetic ablation in the mouse. Dev Dyn. 2004;230:299–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
