

Switching of mesodermal and endodermal properties in hTERT-modified and expanded fetal human pancreatic progenitor cells
- PMID: 20504287
- PMCID: PMC2873697
- DOI: 10.1186/scrt6
Switching of mesodermal and endodermal properties in hTERT-modified and expanded fetal human pancreatic progenitor cells
Erratum in
-
Erratum to: Switching of mesodermal and endodermal properties in hTERT-modified and expanded fetal human pancreatic progenitor cells.Stem Cell Res Ther. 2015 Sep 30;6:189. doi: 10.1186/s13287-015-0176-0. Stem Cell Res Ther. 2015. PMID: 26424142 Free PMC article. No abstract available.
Abstract
Introduction: The ability to expand organ-specific stem/progenitor cells is critical for translational applications, although uncertainties often arise in identifying the lineage of expanded cells. Therefore, superior insights into lineage maintenance mechanisms will be helpful for cell/gene therapy.
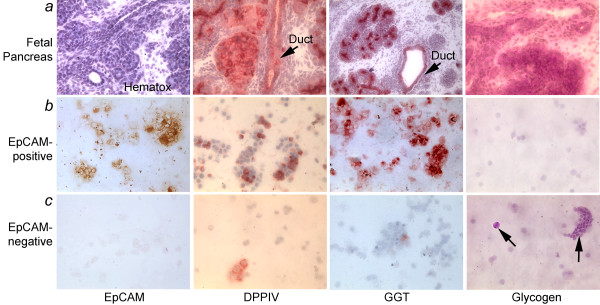
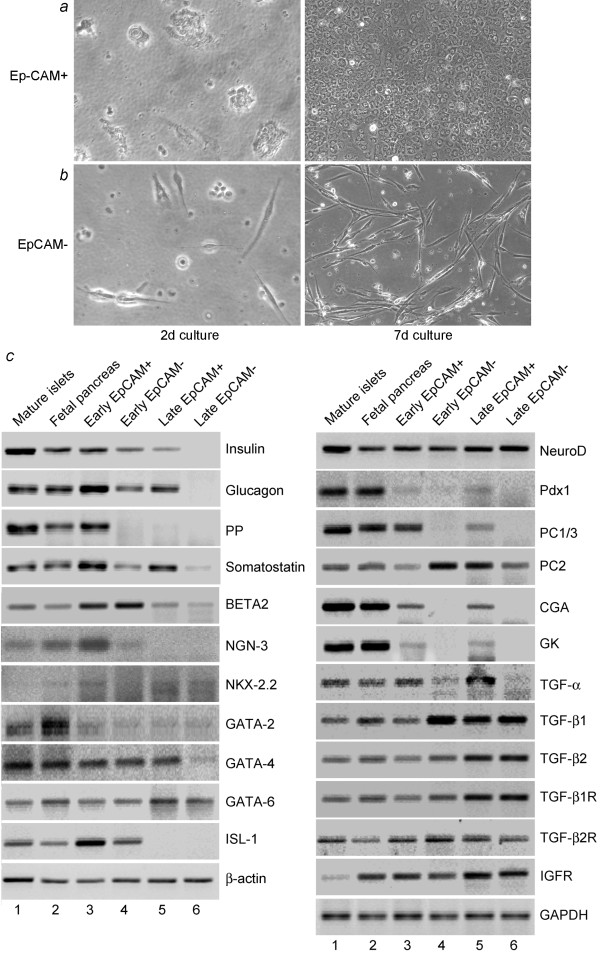
Methods: We studied epithelial cells isolated from fetal human pancreas to assess their proliferation potential, changes in lineage markers during culture, and capacity for generating insulin-expressing beta cells. Cells were isolated by immunomagnetic sorting for epithelial cell adhesion molecule (EpCAM), and characterized for islet-associated transcription factors, hormones, and ductal markers. Further studies were performed after modification of cells with the catalytic subunit of human telomerase reverse transcriptase (hTERT).
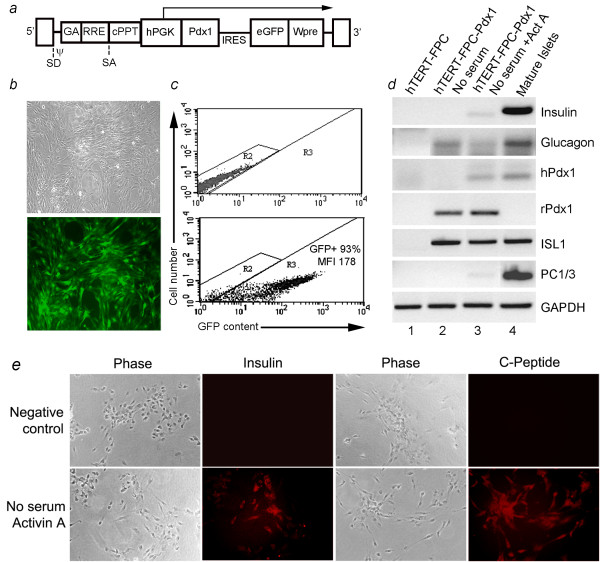
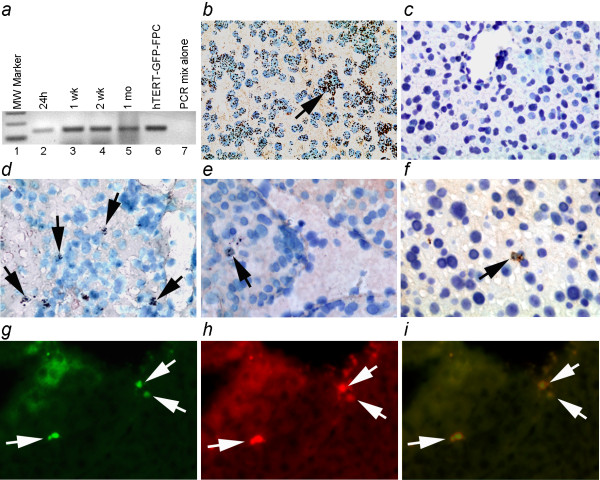
Results: Fetal pancreatic progenitor cells efficiently formed primary cultures, although their replication capacity was limited. This was overcome by introduction and expression of hTERT with a retroviral vector, which greatly enhanced cellular replication in vitro. However, we found that during culture hTERT-modified pancreatic progenitor cells switched their phenotype with gain of additional mesodermal properties. This phenotypic switching was inhibited when a pancreas-duodenal homeobox (Pdx)-1 transgene was expressed in hTERT-modified cells with a lentiviral vector, along with inductive signaling through activin A and serum deprivation. This restored endocrine properties of hTERT-modified cells in vitro. Moreover, transplantation studies in immunodeficient mice verified the capacity of these cells for expressing insulin in vivo.
Conclusions: Limited replication capacity of pancreatic endocrine progenitor cells was overcome by the hTERT mechanism, which should facilitate further studies of such cells, although mechanisms regulating switches between meso-endodermal fates of expanded cells will need to be controlled for developing specific applications. The availability of hTERT-expanded fetal pancreatic endocrine progenitor cells will be helpful for studying and recapitulating stage-specific beta lineage advancement in pluripotent stem cells.
Figures



Similar articles
-
Characterization of an in vitro differentiation assay for pancreatic-like cell development from murine embryonic stem cells: detailed gene expression analysis.Assay Drug Dev Technol. 2011 Aug;9(4):403-19. doi: 10.1089/adt.2010.0314. Epub 2011 Mar 11. Assay Drug Dev Technol. 2011. PMID: 21395400 Free PMC article.
-
Evaluation of islets derived from human fetal pancreatic progenitor cells in diabetes treatment.Stem Cell Res Ther. 2013;4(6):141. doi: 10.1186/scrt352. Stem Cell Res Ther. 2013. PMID: 24268157 Free PMC article.
-
Conversion of immortal liver progenitor cells into pancreatic endocrine progenitor cells by persistent expression of Pdx-1.J Cell Biochem. 2008 May 1;104(1):224-36. doi: 10.1002/jcb.21617. J Cell Biochem. 2008. PMID: 17979180
-
Genes controlling pancreas ontogeny.Int J Dev Biol. 2008;52(7):823-35. doi: 10.1387/ijdb.072444cb. Int J Dev Biol. 2008. PMID: 18956314 Review.
-
Conversion of embryonic stem cells into pancreatic beta-cell surrogates guided by ontogeny.Regen Med. 2006 May;1(3):327-36. doi: 10.2217/17460751.1.3.327. Regen Med. 2006. PMID: 17465786 Review.
Cited by
-
scCATCH: Automatic Annotation on Cell Types of Clusters from Single-Cell RNA Sequencing Data.iScience. 2020 Mar 27;23(3):100882. doi: 10.1016/j.isci.2020.100882. Epub 2020 Feb 4. iScience. 2020. PMID: 32062421 Free PMC article.
-
Pancreatic Stellate Cells: A Rising Translational Physiology Star as a Potential Stem Cell Type for Beta Cell Neogenesis.Front Physiol. 2019 Mar 12;10:218. doi: 10.3389/fphys.2019.00218. eCollection 2019. Front Physiol. 2019. PMID: 30930789 Free PMC article. Review.
-
Biliary tree stem cells, precursors to pancreatic committed progenitors: evidence for possible life-long pancreatic organogenesis.Stem Cells. 2013 Sep;31(9):1966-79. doi: 10.1002/stem.1460. Stem Cells. 2013. PMID: 23847135 Free PMC article.
-
Erratum to: Switching of mesodermal and endodermal properties in hTERT-modified and expanded fetal human pancreatic progenitor cells.Stem Cell Res Ther. 2015 Sep 30;6:189. doi: 10.1186/s13287-015-0176-0. Stem Cell Res Ther. 2015. PMID: 26424142 Free PMC article. No abstract available.
-
The role of fetal pancreatic islet cell transplantation in the treatment of type 2 diabetes mellitus.J Diabetes Metab Disord. 2024 Jun 12;23(2):1949-1957. doi: 10.1007/s40200-024-01448-w. eCollection 2024 Dec. J Diabetes Metab Disord. 2024. PMID: 39610528 Free PMC article.
References
-
- Xu X, D'Hoker J, Stangé G, Bonné S, De Leu N, Xiao X, Casteele M Van de, Mellitzer G, Ling Z, Pipeleers D, Bouwens L, Scharfmann R, Gradwohl G, Heimberg H. β Cells Can Be Generated from Endogenous Progenitors in Injured Adult Mouse Pancreas. Cell. 2008;132:197–207. doi: 10.1016/j.cell.2007.12.015. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
