

doi: 10.1093/nar/gkq437.
Epub 2010 May 26.
Northern blot detection of endogenous small RNAs (approximately14 nt) in bacterial total RNA extracts
Affiliations
- PMID: 20504856
- PMCID: PMC2919735
- DOI: 10.1093/nar/gkq437
Item in Clipboard
Northern blot detection of endogenous small RNAs (approximately14 nt) in bacterial total RNA extracts
Nucleic Acids Res.
2010 Aug.
Abstract
Here we describe a northern blot procedure that allows the detection of endogenous RNAs as small as approximately 14 nt in total RNA extracts from bacteria. RNAs that small and as part of total bacterial RNA extracts usually escape detection by northern blotting. The approach combines LNA probes 5'-digoxigenin-endlabeled for non-radioactive probe detection with 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide-mediated chemical crosslinking of RNAs to nylon membranes, and necessitates the use of native PAGE either with the TBE or MOPS buffer system.
Figures
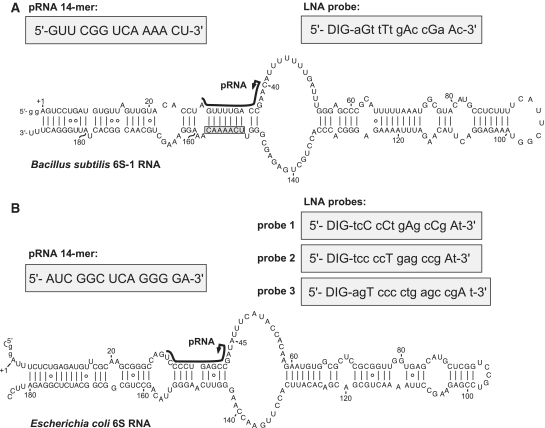
Secondary structure presentation of (A) mature B. subtilis 6S-1 RNA (190 nt), adapted from (11), and (B) of E. coli 6S RNA according to (9); both RNAs were transcribed with two artificially added G residues (lowercase letters) for reasons of efficient synthesis by T7 RNA polymerase. In both panels, the black lines along the sequence indicate the region of B. subtilis 6S-1 RNA and E. coli 6S RNA that serve as a template for the synthesis of small transcripts (product RNAs = pRNAs) by RNA polymerase (this study, 9); arrows mark the starting point of pRNA transcription; the chemically synthesized pRNA mimics are depicted in the grey boxes on the left above the 6S RNA structures. In panel A, 6S-1 RNA nt 151–157 complementary to nt 2–8 of the LNA probe are boxed. LNA probes for pRNA detection are shown in grey boxes on the right above the secondary structures; DIG, digoxigenin attached to the 5′-end via a C6 linker; small letters depict DNA residues, capital letters LNA residues.
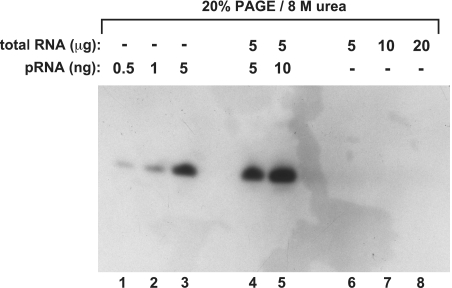
Standard denaturing northern blot using the classical ‘baking’ of the membrane at 80°C for RNA immobilization. Lanes 1–3: various amounts of the chemically synthesized pRNA 14-mer (with 5′-OH terminus; Figure 1A); lanes 4 and 5: 5 or 10 ng of the chemically synthesized pRNA 14-mer mixed with 5 µg of total RNA from B. subtilis before electrophoresis and membrane transfer; lanes 6–8: samples with varying amounts of total RNA from B. subtilis for the purpose of endogenous pRNA detection. Total cellular RNA was isolated 3 min after outgrowth from stationary phase using the hot phenol method; the 5′-digoxigenin-labeled LNA–DNA mixmer shown in Figure 1A was used as probe. (for details, see ‘Materials and Methods’ section).
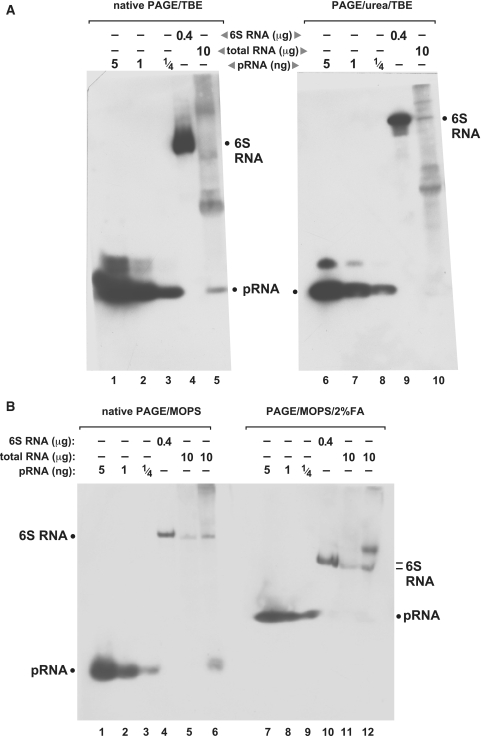
Comparison of northern blots performed in different buffer systems under native and denaturing conditions, using EDC crosslinking for RNA immobilization on nylon membranes. (A) Northern blots of RNA separated by native (left) and denaturing (right) PAGE using the TBE buffer system. Lanes 1–3 and 6–8: varying amounts of the chemically synthesized pRNA 14-mer (Figure 1A); lanes 4 and 9: 400 ng of in vitro transcribed B. subtilis 6S-1 RNA loaded onto the gels; the northern blot signal is attributable to the probe’s complementarity to seven consecutive nucleotides of 6S-1 RNA (nts 151–157; Figure 1A), including three LNA/RNA base pairs; lanes 5 and 10: 10 µg of total cellular RNA prepared from B. subtilis cells after 3 min of outgrowth from stationary phase (‘Materials and Methods’ section); the endogenous pRNA yielded a signal under native but not under denaturing conditions (lanes 5 versus 10). Likewise, pRNA signals in lanes 1–3 were more intense than those in lanes 6–8. (B) The same samples as in panel A were analyzed by MOPS-buffered PAGE and northern blotting in the absence (lanes 1–6) and presence (lanes 7–12) of 2% formaldehyde as denaturing agent. Additionally, total RNA prepared from stationary phase cells was loaded onto the gels (lanes 5 and 11); endogenous pRNA in the outgrowth RNA fraction was again only detectable under native conditions (compare lanes 6 and 12). Note that PAGE with MOPS and formaldehyde generally resulted in less efficient separation of RNAs according to size. Film exposition times were 20–45 min; the two part figures in panel A originate from the same blot and exposed film; exposition time in panel B slightly differed from that in panel A; criterion for exposition length was good visibility of the pRNA signal (lane 5 in panel A, lane 6 in panel B), thus often two or more films were exposed to the same blot for different periods of time to optimize blot performance.
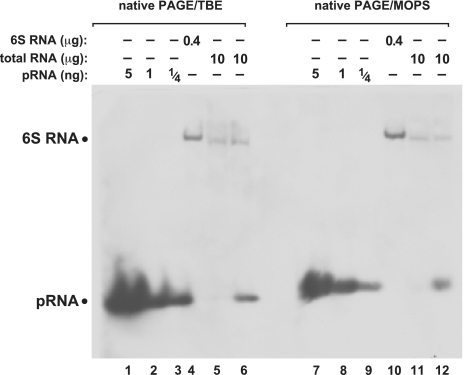
Comparison of native PAGE northern blots using the TBE (lanes 1–6) or MOPS (lanes 7–12) buffer systems. The two different gels were blotted simultaneously onto the same nylon membrane. For further details, see legend to Figure 3B.
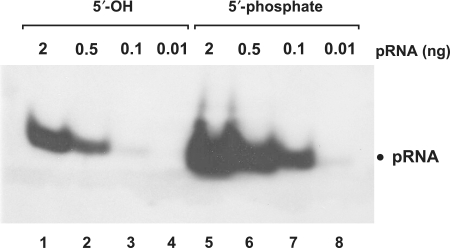
Functional groups at the 5′-terminus have a strong impact on signal strength. pRNA with a 5′-phosphate (lanes 5–8) yielded enhanced signal strength (∼10-fold) compared to the same RNA with a 5′-hydroxyl end (lanes 1–4). Gel electrophoresis: 10% native PAGE, 1× TBE; RNAs were coupled to nylon membranes by EDC crosslinking.
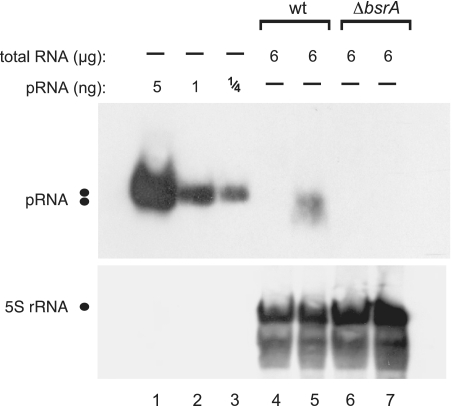
Specificity of pRNA detection demonstrated by using total RNA from the B. subtilis wild-type (wt) strain PY79 (lanes 4 and 5) versus a 6S-1 RNA knockout derivative strain, PY79 ΔbsrA (lanes 6 and 7); lanes 4 and 6, total RNA from cells harvested in stationary phase; lanes 5 and 7, total RNA from cells harvested during outgrowth (for details, see ‘Materials and Methods’ section). 5S rRNA was used as an internal loading control in lanes 4–7; since we used native PAGE here, the expanded area of 5S rRNA signals may be due to different 5S rRNA conformers.
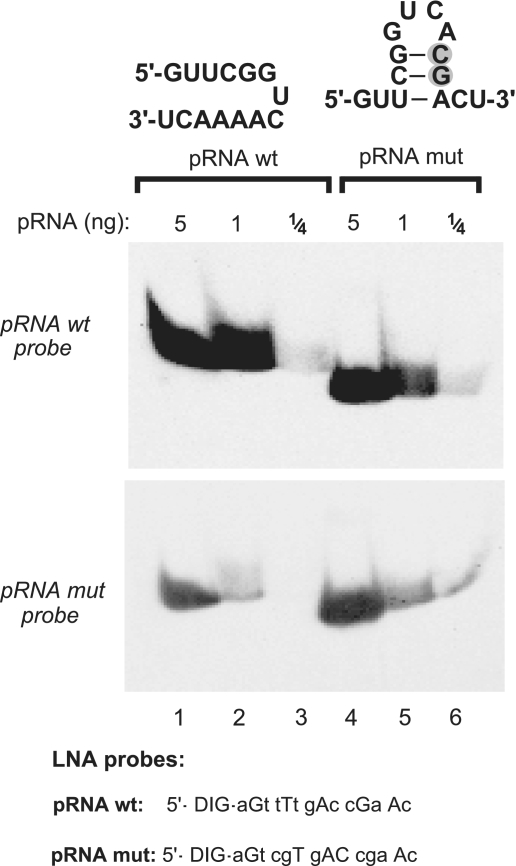
Evaluation of the influence of potential intramolecular RNA secondary structures on detection efficiency. A mutated version of the 6S-1 pRNA (pRNA mut with two consecutive point mutations, grey-shaded; lanes 4–6), designed to have the potential to form a hairpin with a 3-bp stem, migrated somewhat faster than wt pRNA (lanes 4–6 versus 1–3) in a 20% native PAA gel (TBE buffer system), suggesting that the hairpin indeed formed at least transiently during electrophoresis. The two pRNA variants (carrying 5′-OH termini) were mutually detected with LNA probes either complementary to wt pRNA (top panel) or to pRNA mut (bottom panel), with LNA probes specified below the blots (uppercase letters indicate LNA and lowercase letters DNA residues). Films were exposed to the blot for 35 min.
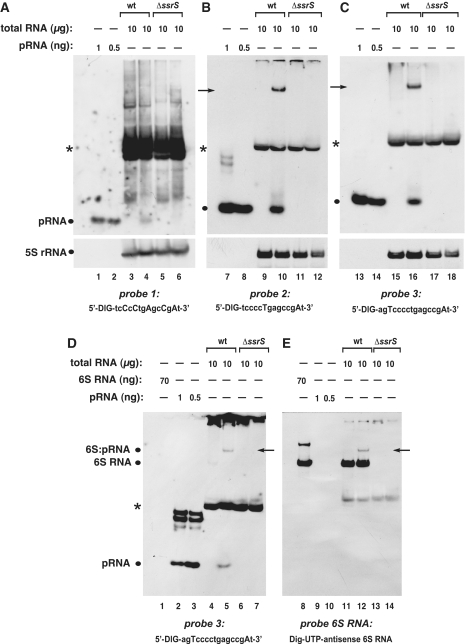
Specificity of different probes for the detection of pRNAs in RNA extracts from E. coli. (A–C) Detection of pRNAs derived from E. coli 6S RNA in total RNA extracts from the wild-type (wt) strain MG1655 (lanes 3, 4, 9, 10, 15 and 16) versus RNA extracts derived from a MG1655 6S RNA knockout (ΔssrS) strain (lanes 5, 6, 11, 12, 17 and 18) using three different probes shown below each panel (uppercase letters indicate LNA and lowercase letters DNA residues). Deviating from our standard procedure (‘Materials and Methods’ section), RNA samples were heated in denaturing loading buffer for 5 min at 95°C immediately before gel loading. In analogy to B. subtilis (Figure 6), RNA harvested from cells in stationary phase (lanes 3, 5, 9, 11, 15 and 17) or during outgrowth (lanes 4, 6, 10, 12, 16 and 18) was analyzed. The chemically synthesized E. coli pRNA 14-mer (Figure 1B) was loaded as size control in lanes 1, 2, 7, 8, 13 and 14 (pRNA positions marked by dots at the left margin of each panel). NC filters were stripped after hybridization with pRNA probes and hybridized with a 5S rRNA-specific probe (as loading control) shown at the bottom of each panel. A pRNA signal was seen under outgrowth conditions for wt bacteria (lanes 4, 10 and 16), but not for ΔssrS bacteria (lanes 6, 12, 18). Probe 1 (A) was the least specific and yielded the highest background among the three different probes, which we attribute to the high number of five LNA residues, two of which are in the tetracytidylate stretch that is able to basepair with tetraguanylates. Signals indicated by arrows in lanes 10 and 16 are 6S RNA:pRNA hybrids (see below); asterisks mark a cross-hybridizing RNA of unknown identity. (D and E) The two panels are halves of the same blot, representing identical sample sets, but hybridized to different probes (probe 3 in D, antisense 6S RNA in E). The signals in lanes 5 and 12, indicated by arrows, are inferred to be 6S RNA:pRNA hybrids, as they migrate at identical position and are detected with both probes. For control, in vitro transcribed E. coli 6S RNA was loaded (lanes 1 and 8). All samples in D and E were heated for 2 min at 95°C in denaturing loading buffer immediately before gel loading. Deviating from the standard protocol (‘Materials and Methods’ section), the hybridization temperature was 72°C instead of 50°C for probe 1, and 68°C instead of 50°C for probes 2 and 3.
References
-
- Várallyay E, Burgyán J, Havelda Z. Detection of microRNAs by northern blot analyses using LNA probes. Methods. 2007;43:140–145. - PubMed
-
- Berezikov E, Thuemmler F, van Laake LW, Kondova I, Bontrop R, Cuppen E, Plasterk RHA. Diversity of microRNAs in human and chimpanzee brain. Nat. Genet. 2006;38:1375–1377. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
