

SUMO functions in constitutive transcription and during activation of inducible genes in yeast
- PMID: 20504900
- PMCID: PMC2885660
- DOI: 10.1101/gad.1917910
SUMO functions in constitutive transcription and during activation of inducible genes in yeast
Abstract
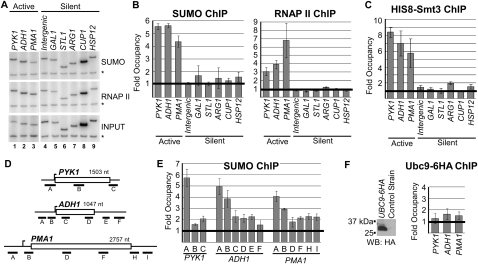
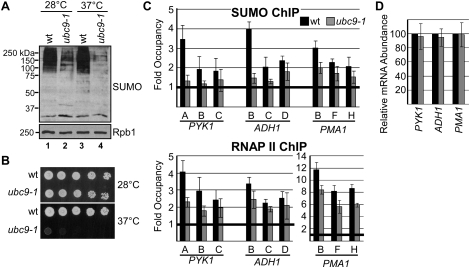
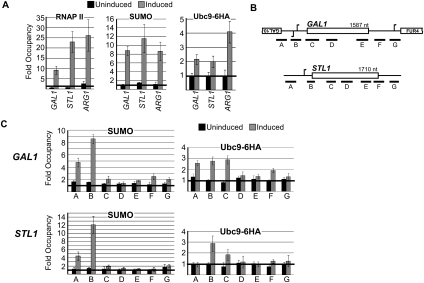
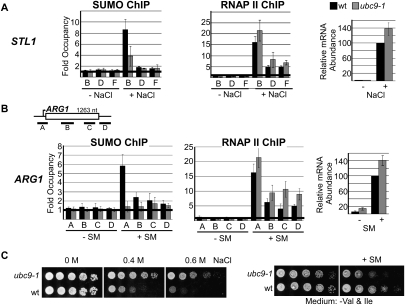
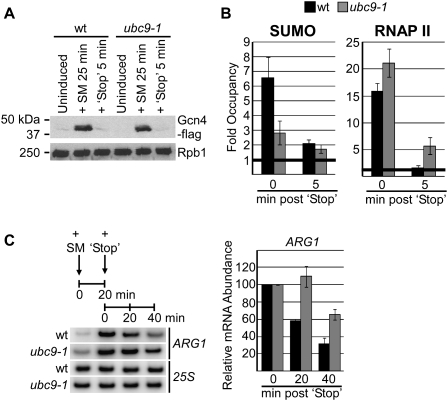
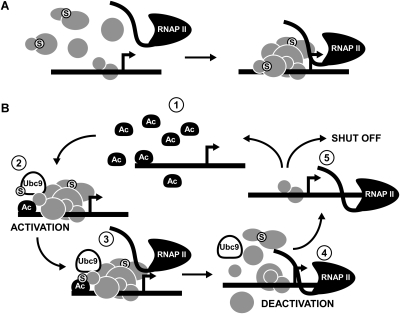
Transcription factors represent one of the largest groups of proteins regulated by SUMO (small ubiquitin-like modifier) modification, and their sumoylation is usually associated with transcriptional repression. To investigate whether sumoylation plays a general role in regulating transcription in yeast, we determined the occupancy of sumoylated proteins at a variety of genes by chromatin immunoprecipitation (ChIP) using an antibody that recognizes the yeast SUMO peptide. Surprisingly, we detected sumoylated proteins at all constitutively transcribed genes tested but not at repressed genes. Ubc9, the SUMO conjugation enzyme, was not present on these genes, but its inactivation reduced SUMO at the constitutive promoters and modestly decreased RNA polymerase II levels. In contrast, activation of the inducible GAL1, STL1, and ARG1 genes caused not only a striking accumulation of SUMO at all three promoter regions, but also recruitment of Ubc9, indicating that gene activation involves sumoylation of promoter-bound factors. However, Ubc9 inactivation, while reducing sumoylation at the induced promoters, paradoxically resulted in increased transcription. Providing an explanation for this, the reduced sumoylation impaired the cell's ability to appropriately shut off transcription of the induced ARG1 gene, indicating that SUMO can facilitate transcriptional silencing. Our findings thus establish unexpected roles for sumoylation in both constitutive and activated transcription, and provide a novel mechanism for regulating gene expression.
Figures
References
-
- Auld KL, Brown CR, Casolari JM, Komili S, Silver PA 2006. Genomic association of the proteasome demonstrates overlapping gene regulatory activity with transcription factor substrates. Mol Cell 21: 861–871 - PubMed
-
- Betting J, Seufert W 1996. A yeast Ubc9 mutant protein with temperature-sensitive in vivo function is subject to conditional proteolysis by a ubiquitin- and proteasome-dependent pathway. J Biol Chem 271: 25790–25796 - PubMed
-
- Bhaumik SR, Malik S 2008. Diverse regulatory mechanisms of eukaryotic transcriptional activation by the proteasome complex. Crit Rev Biochem Mol Biol 43: 419–433 - PubMed
-
- Boyer-Guittaut M, Birsoy K, Potel C, Elliott G, Jaffray E, Desterro JM, Hay RT, Oelgeschlager T 2005. SUMO-1 modification of human transcription factor (TF) IID complex subunits: Inhibition of TFIID promoter-binding activity through SUMO-1 modification of hsTAF5. J Biol Chem 280: 9937–9945 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous