

Pregnancy and the endocrine regulation of the baroreceptor reflex
- PMID: 20504907
- PMCID: PMC2928618
- DOI: 10.1152/ajpregu.00059.2010
Pregnancy and the endocrine regulation of the baroreceptor reflex
Abstract
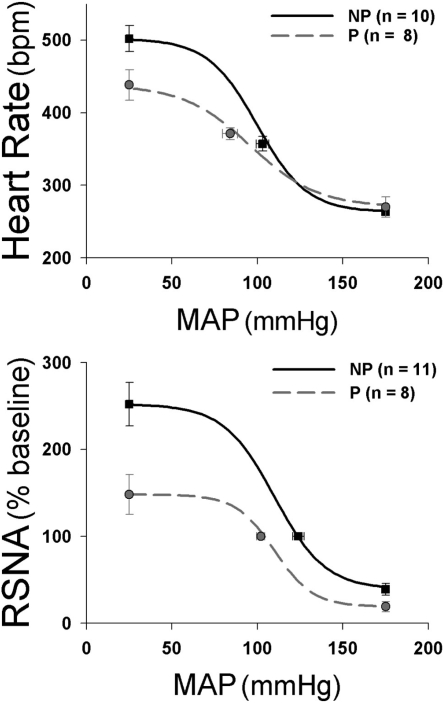
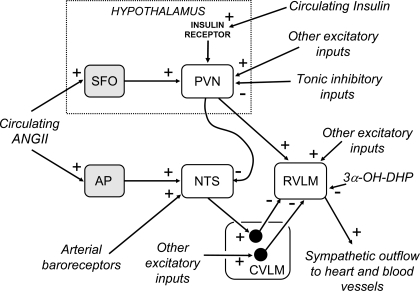
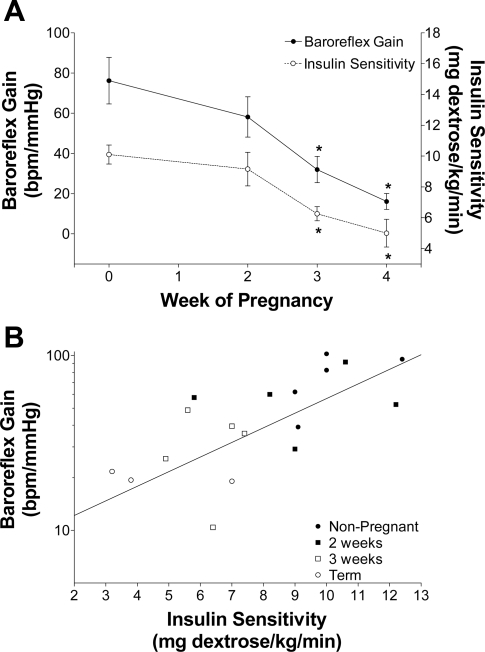
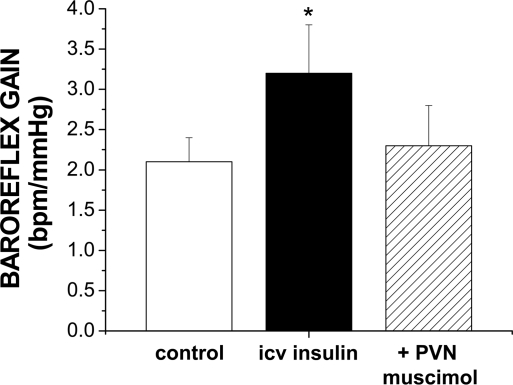
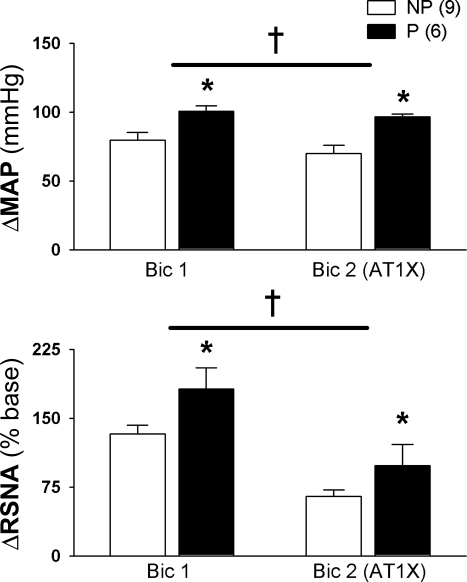
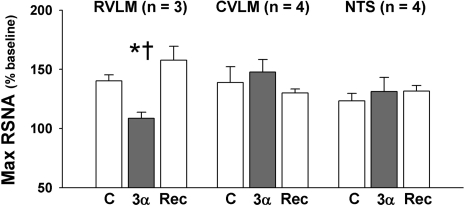
The purpose of this review is to delineate the general features of endocrine regulation of the baroreceptor reflex, as well as specific contributions during pregnancy. In contrast to the programmed changes in baroreflex function that occur in situations initiated by central command (e.g., exercise or stress), the complex endocrine milieu often associated with physiological and pathophysiological states can influence the central baroreflex neuronal circuitry via multiple sites and mechanisms, thereby producing varied changes in baroreflex function. During pregnancy, baroreflex gain is markedly attenuated, and at least two hormonal mechanisms contribute, each at different brain sites: increased levels of the neurosteroid 3alpha-hydroxy-dihydroprogesterone (3alpha-OH-DHP), acting in the rostral ventrolateral medulla (RVLM), and reduced actions of insulin in the forebrain. 3alpha-OH-DHP appears to potentiate baroreflex-independent GABAergic inhibition of premotor neurons in the RVLM, which decreases the range of sympathetic nerve activity that can be elicited by changes in arterial pressure. In contrast, reductions in the levels or actions of insulin in the brain blunt baroreflex efferent responses to increments or decrements in arterial pressure. Although plasma levels of angiotensin II are increased in pregnancy, this is not responsible for the reduction in baroreflex gain, although it may contribute to the increased level of sympathetic nerve activity in this condition. How these different hormonal effects are integrated within the brain, as well as possible interactions with additional potential neuromodulators that influence baroreflex function during pregnancy and other physiological and pathophysiological states, remains to be clearly delineated.
Figures
Similar articles
-
Neurosteroid modulation of arterial baroreflex function in the rostral ventrolateral medulla.Auton Neurosci. 2011 Apr 26;161(1-2):28-33. doi: 10.1016/j.autneu.2010.10.002. Epub 2010 Nov 10. Auton Neurosci. 2011. PMID: 21071286
-
Angiotensin peptides and baroreflex control of sympathetic outflow: pathways and mechanisms of the medulla oblongata.Brain Res Bull. 2000 Jan 15;51(2):119-28. doi: 10.1016/s0361-9230(99)00237-3. Brain Res Bull. 2000. PMID: 10709957 Review.
-
Galanin microinjection into rostral ventrolateral medulla of the rat is hypotensive and attenuates sympathetic chemoreflex.Am J Physiol Regul Integr Comp Physiol. 2009 Apr;296(4):R1019-26. doi: 10.1152/ajpregu.90885.2008. Epub 2009 Jan 21. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19158406
-
Neurosteroid modulation of arterial baroreflex-sensitive neurons in rat rostral ventrolateral medulla.Am J Physiol. 1998 Apr;274(4):R903-11. doi: 10.1152/ajpregu.1998.274.4.R903. Am J Physiol. 1998. PMID: 9575950
-
Regulation of sympathetic nervous system function after cardiovascular deconditioning.Ann N Y Acad Sci. 2001 Jun;940:454-68. doi: 10.1111/j.1749-6632.2001.tb03698.x. Ann N Y Acad Sci. 2001. PMID: 11458701 Review.
Cited by
-
Association between temporal patterns of baroreflex sensitivity after traumatic brain injury and prognosis: a preliminary study.Neurol Sci. 2023 May;44(5):1653-1663. doi: 10.1007/s10072-022-06579-7. Epub 2023 Jan 7. Neurol Sci. 2023. PMID: 36609622 Free PMC article.
-
Impaired baroreflex gain during pregnancy in conscious rats: role of brain insulin.Hypertension. 2011 Feb;57(2):283-8. doi: 10.1161/HYPERTENSIONAHA.110.162354. Epub 2010 Dec 13. Hypertension. 2011. PMID: 21149828 Free PMC article.
-
Posture-related changes in sympathetic baroreflex sensitivity during normal pregnancy.Clin Auton Res. 2022 Dec;32(6):485-495. doi: 10.1007/s10286-022-00903-z. Epub 2022 Nov 17. Clin Auton Res. 2022. PMID: 36394777 Free PMC article.
-
Adaptations in autonomic nervous system regulation in normal and hypertensive pregnancy.Handb Clin Neurol. 2020;171:57-84. doi: 10.1016/B978-0-444-64239-4.00003-5. Handb Clin Neurol. 2020. PMID: 32736759 Free PMC article. Review.
-
Pregnancy in a woman with a Fontan circulation: A review.Obstet Med. 2018 Mar;11(1):6-11. doi: 10.1177/1753495X17737680. Epub 2017 Nov 22. Obstet Med. 2018. PMID: 29636807 Free PMC article. Review.
References
-
- Andresen MC, Doyle MW, Jin YH, Bailey TW. Cellular mechanisms of baroreceptor integration at the nucleus tractus solitarius. Ann NY Acad Sci 940: 132–141, 2001 - PubMed
-
- Anglin JC, Brooks VL. Tyrosine hydroxylase and norepinephrine transporter in sympathetic ganglia of female rats vary with reproductive state. Auton Neurosci 105: 8–15, 2003 - PubMed
-
- Avery ND, Wolfe LA, Amara CE, Davies GA, McGrath MJ. Effects of human pregnancy on cardiac autonomic function above and below the ventilatory threshold. J Appl Physiol 90: 321–328, 2001 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources