

The activity of hyaluronan synthase 2 is regulated by dimerization and ubiquitination
- PMID: 20507985
- PMCID: PMC2911313
- DOI: 10.1074/jbc.M110.127050
The activity of hyaluronan synthase 2 is regulated by dimerization and ubiquitination
Abstract
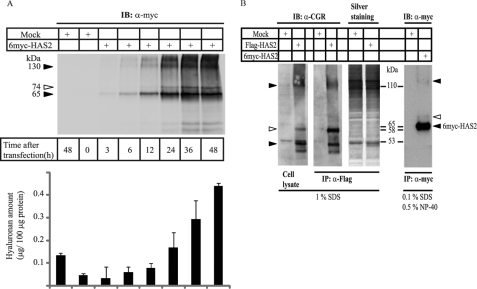
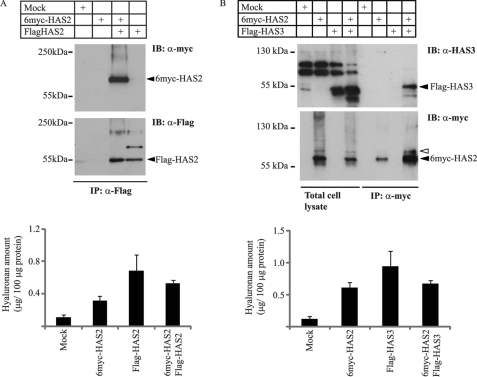
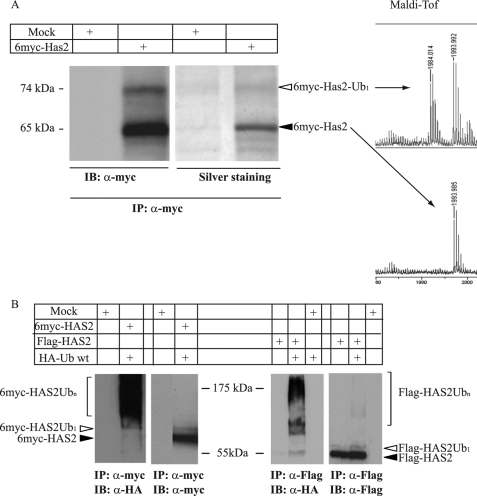
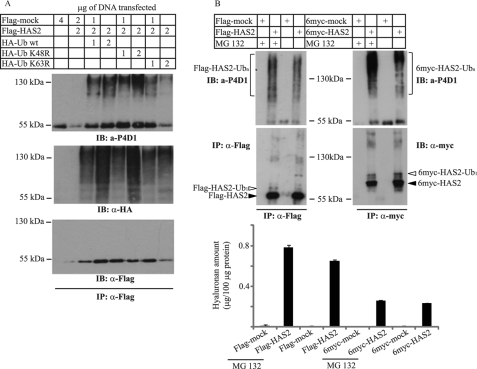
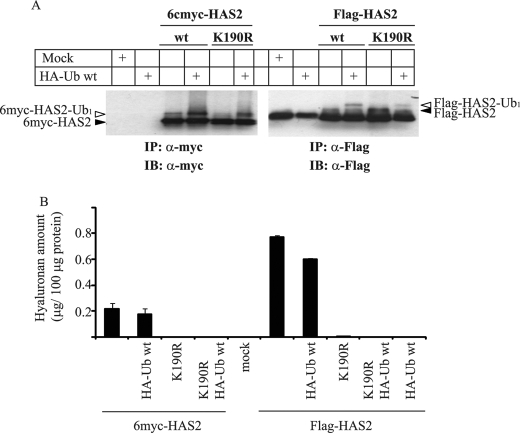
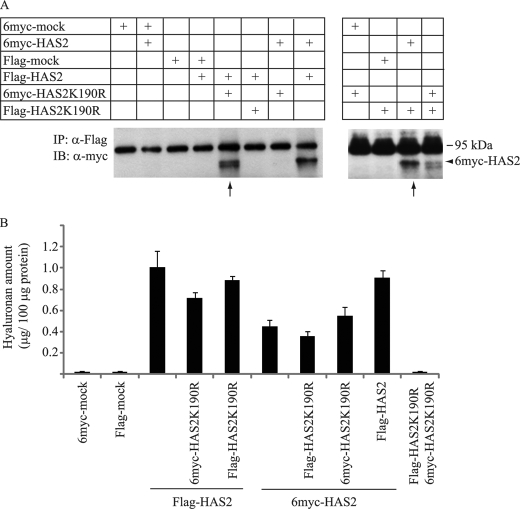
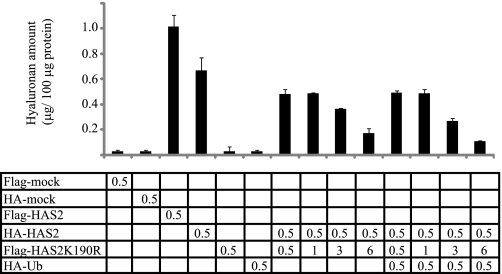
Hyaluronan is a component of the extracellular matrix, which affects tissue homeostasis. In this study, we investigated the regulatory mechanisms of one of the hyaluronan-synthesizing enzymes, HAS2. Ectopic expression of Flag- and 6myc-HAS2 in COS-1 cells followed by immunoprecipitation and immunoblotting revealed homodimers; after co-transfection with Flag-HAS3, also heterodimers were seen. Furthermore, the expressed HAS2 was ubiquitinated. We identified one acceptor site for ubiquitin on lysine residue 190. Mutation of this residue led to inactivation of the enzymatic activity of HAS2. Interestingly, K190R-mutated HAS2 formed dimers with wt HAS2 and quenched the activity of wt HAS2, thus demonstrating a functional role of the dimeric configuration.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
