

Mechanical signals control SOX-9, VEGF, and c-Myc expression and cell proliferation during inflammation via integrin-linked kinase, B-Raf, and ERK1/2-dependent signaling in articular chondrocytes
- PMID: 20509944
- PMCID: PMC2911896
- DOI: 10.1186/ar3039
Mechanical signals control SOX-9, VEGF, and c-Myc expression and cell proliferation during inflammation via integrin-linked kinase, B-Raf, and ERK1/2-dependent signaling in articular chondrocytes
Abstract
Introduction: The importance of mechanical signals in normal and inflamed cartilage is well established. Chondrocytes respond to changes in the levels of proinflammatory cytokines and mechanical signals during inflammation. Cytokines like interleukin (IL)-1beta suppress homeostatic mechanisms and inhibit cartilage repair and cell proliferation. However, matrix synthesis and chondrocyte (AC) proliferation are upregulated by the physiological levels of mechanical forces. In this study, we investigated intracellular mechanisms underlying reparative actions of mechanical signals during inflammation.
Methods: ACs isolated from articular cartilage were exposed to low/physiologic levels of dynamic strain in the presence of IL-1beta. The cell extracts were probed for differential activation/inhibition of the extracellular signal-regulated kinase 1/2 (ERK1/2) signaling cascade. The regulation of gene transcription was examined by real-time polymerase chain reaction.
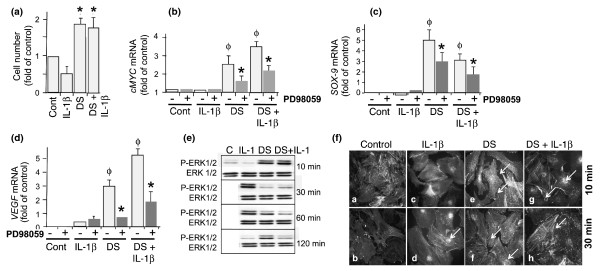
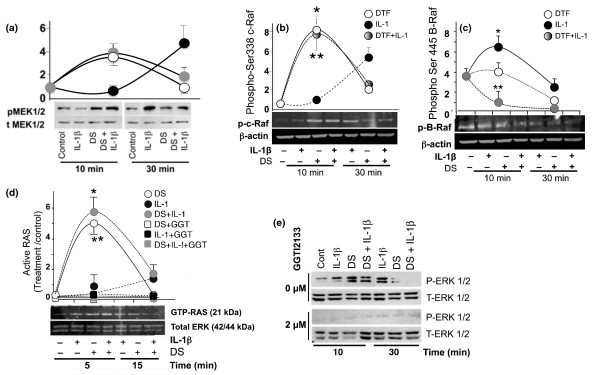
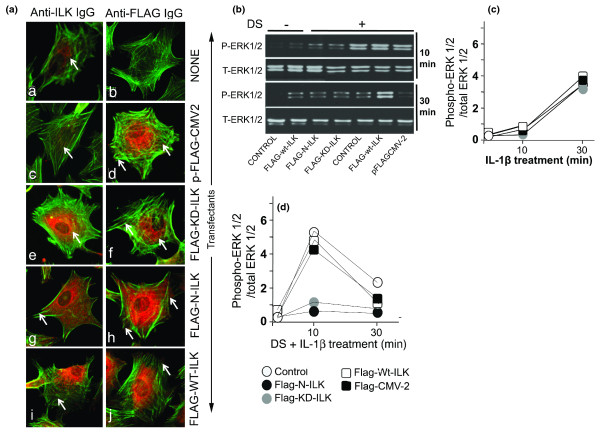
Results: Mechanoactivation, but not IL-1beta treatment, of ACs initiated integrin-linked kinase activation. Mechanical signals induced activation and subsequent C-Raf-mediated activation of MAP kinases (MEK1/2). However, IL-1beta activated B-Raf kinase activity. Dynamic strain did not induce B-Raf activation but instead inhibited IL-1beta-induced B-Raf activation. Both mechanical signals and IL-1beta induced ERK1/2 phosphorylation but discrete gene expression. ERK1/2 activation by mechanical forces induced SRY-related protein-9 (SOX-9), vascular endothelial cell growth factor (VEGF), and c-Myc mRNA expression and AC proliferation. However, IL-1beta did not induce SOX-9, VEGF, and c-Myc gene expression and inhibited AC cell proliferation. More importantly, SOX-9, VEGF, and Myc gene transcription and AC proliferation induced by mechanical signals were sustained in the presence of IL-1beta.
Conclusions: The findings suggest that mechanical signals may sustain their effects in proinflammatory environments by regulating key molecules in the MAP kinase signaling cascade. Furthermore, the findings point to the potential of mechanosignaling in cartilage repair during inflammation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
