

Calcium clearance and its energy requirements in cerebellar neurons
- PMID: 20510449
- PMCID: PMC2900537
- DOI: 10.1016/j.ceca.2010.04.004
Calcium clearance and its energy requirements in cerebellar neurons
Abstract
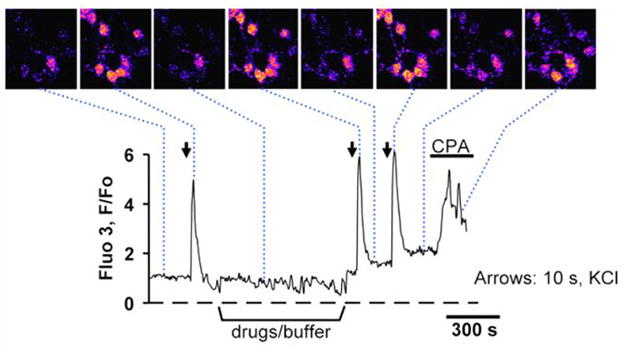
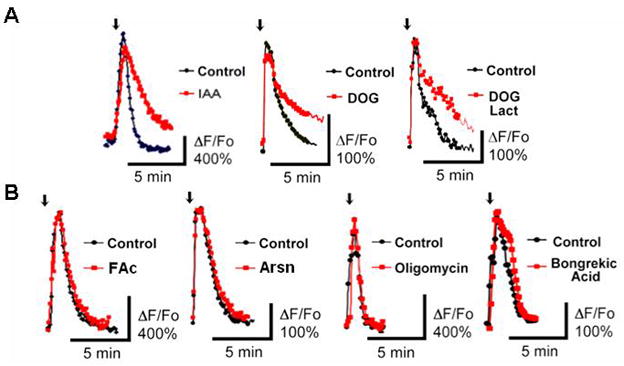
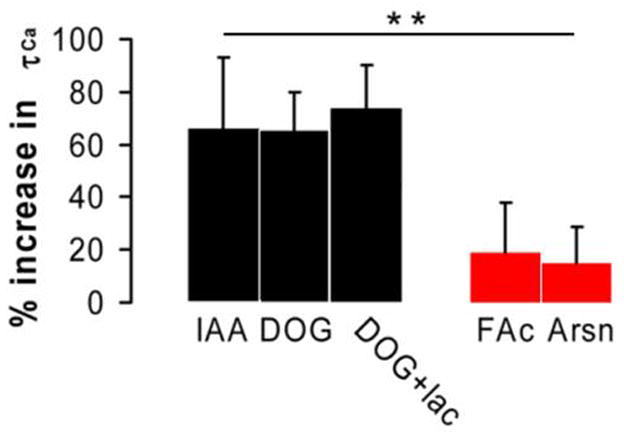
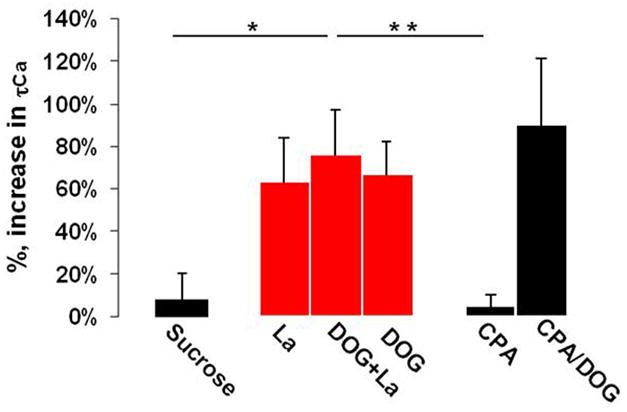
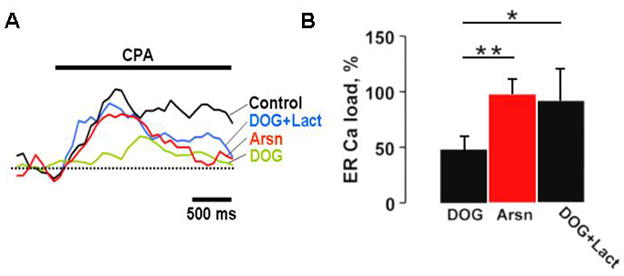
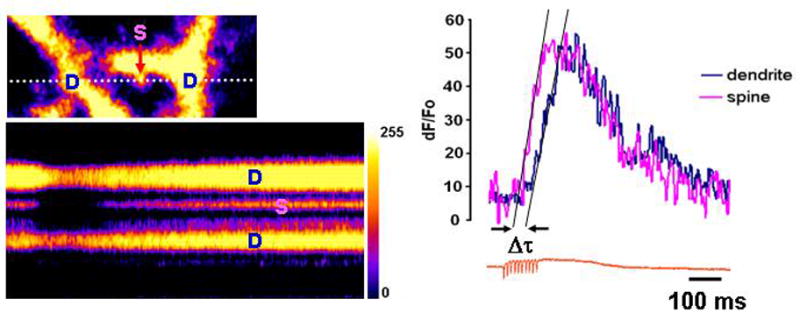
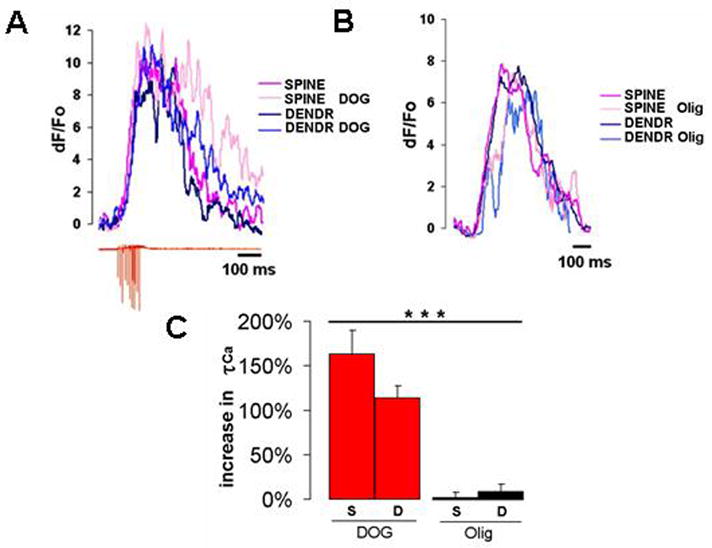
Quick cytosolic calcium clearance is essential for the effective modulation of various cellular functions. An excess of cytosolic calcium after influx is largely removed via ATP-dependent mechanisms located in the plasma membrane and the endoplasmic reticulum. Therefore, calcium clearance depends critically on the adequate supply of ATP, which may come from either glycolysis or mitochondria, or both. However, it presently remains unknown the degree to which individual ATP generating pathways - glycolysis and mitochondria power ATP-dependent calcium as well as other vital ion clearance mechanisms in neurons. In this study, we explored the relationship between the energy generating pathways and ion clearance mechanisms in neurons by characterizing the effects of glycolytic and mitochondrial inhibitors of ATP synthesis on calcium clearance kinetics in the soma, dendrites and spines. Stimulation of cultured cerebellar granule cells by brief pulses of 60mM potassium ACSF, and electrical stimulation of purkinje cells in acutely prepared slices led to a transient calcium influx, whose clearance was largely mediated by the plasma membrane Ca(2+)-ATPase pump. Inhibition of glycolysis by deoxyglucose or iodoacetic acid resulted in a marked slowing in calcium clearance in the soma, dendrites, and spines with the spines affected the most. However, inhibition of the mitochondrial citric acid cycle with fluoroacetate and arsenite, or mitochondrial ATP synthase with oligomycin did not produce any immediate effects on calcium clearance kinetics in any of those neuronal regions. Although cytosolic calcium clearance was not affected by the inhibition of mitochondria, the magnitude of the calcium clearance delay induced by glycolytic inhibitors in different neuronal compartments was related to their mitochondrial density. Conversely, the endoplasmic reticulum Ca(2+)-ATPase pump activity is fuelled by both glycolytic and mitochondrial ATP, as evidenced by a minimal change in the endoplasmic reticulum calcium contents in cells treated with either deoxyglucose supplemented with lactate or arsenite. Taken together, these data suggest that calcium clearance in cerebellar granule and purkinje cells relies on the plasma membrane Ca(2+)-ATPase, and is powered by glycolysis.
2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Matsumura F, Clark JM. ATP-utilizing systems in the squid axons: a review on the biochemical aspects of ion-transport. Prog Neurobiol. 1982;18:231–55. - PubMed
-
- Atwell D, Laughlin SB. An energy budget for signaling in the grey matter of the brain. J Cereb Blood Flow Metab. 2001;21:1133–1145. - PubMed
-
- Lenny P. The cost of cortical computation. Curr Biol. 2003;13:493–7. - PubMed
-
- Bano D, Young KW, Guerin CJ, Lefeuvre R, Rothwell NJ, Naldini L, Rizzuto R, Carafoli E, Nicotera P. Cleavage of the plasma membrane Na+/Ca2+ exchanger in excitotoxicity. Cell. 2005;120:275–285. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
