

AID and somatic hypermutation
- PMID: 20510733
- PMCID: PMC2954419
- DOI: 10.1016/S0065-2776(10)05006-6
AID and somatic hypermutation
Abstract
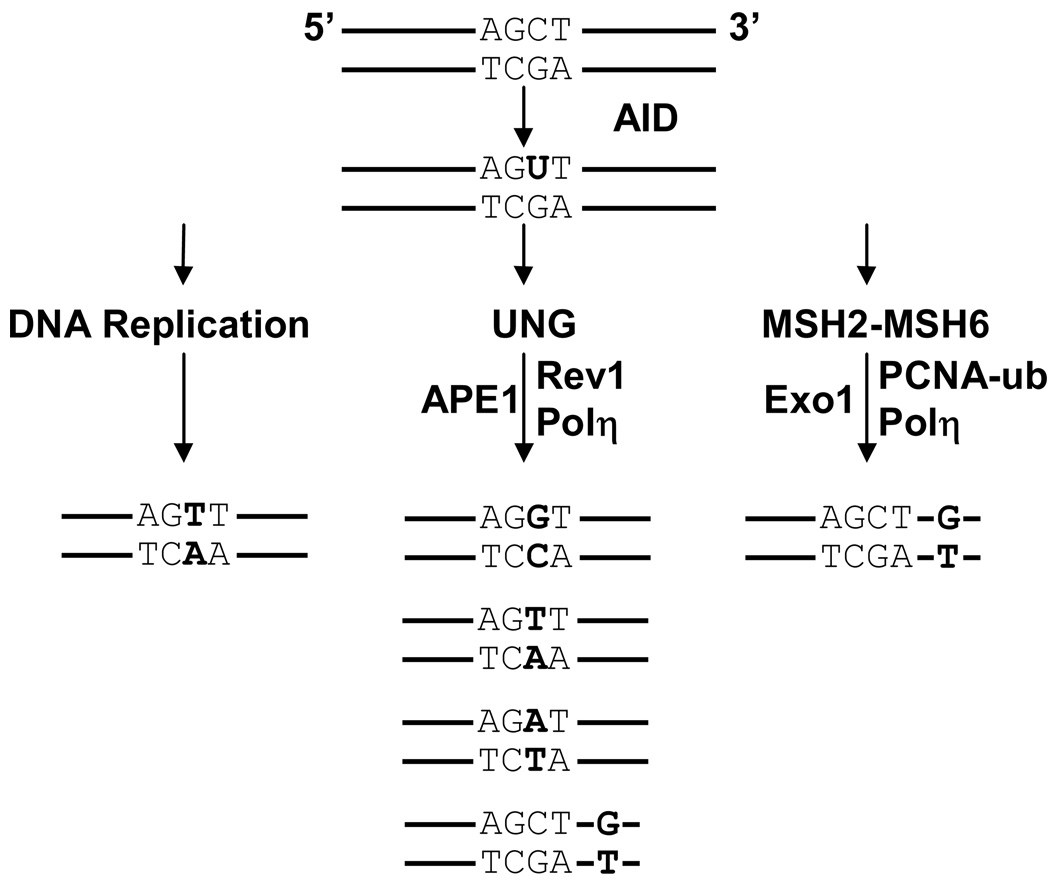
In response to an assault by foreign organisms, peripheral B cells can change their antibody affinity and isotype by somatically mutating their genomic DNA. The ability of a cell to modify its DNA is exceptional in light of the potential consequences of genetic alterations to cause human disease and cancer. Thus, as expected, this mechanism of antibody diversity is tightly regulated and coordinated through one protein, activation-induced deaminase (AID). AID produces diversity by converting cytosine to uracil within the immunoglobulin loci. The deoxyuracil residue is mutagenic when paired with deoxyguanosine, since it mimics thymidine during DNA replication. Additionally, B cells can manipulate the DNA repair pathways so that deoxyuracils are not faithfully repaired. Therefore, an intricate balance exists which is regulated at multiple stages to promote mutation of immunoglobulin genes, while retaining integrity of the rest of the genome. Here we discuss and summarize the current understanding of how AID functions to cause somatic hypermutation.
(c) 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Arakawa H, Hauschild J, Buerstedde JM. Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion. Science. 2002;295:1301–1306. - PubMed
-
- Bachl J, Carlson C, Gray-Schopfer V, Dessing M, Olsson C. Increased transcription levels induce higher mutation rates in a hypermutating cell line. J Immunol. 2001;166:5051–5057. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
