

Deactivation of L-type Ca current by inhibition controls LTP at excitatory synapses in the cerebellar nuclei
- PMID: 20510859
- PMCID: PMC2886803
- DOI: 10.1016/j.neuron.2010.04.024
Deactivation of L-type Ca current by inhibition controls LTP at excitatory synapses in the cerebellar nuclei
Abstract
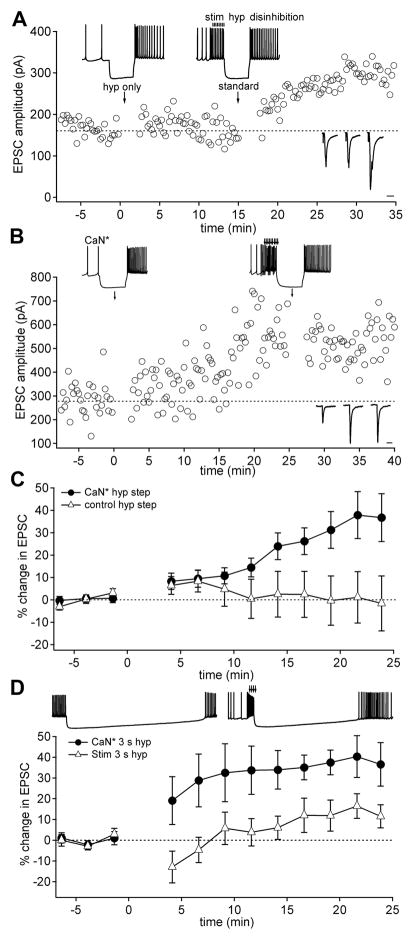
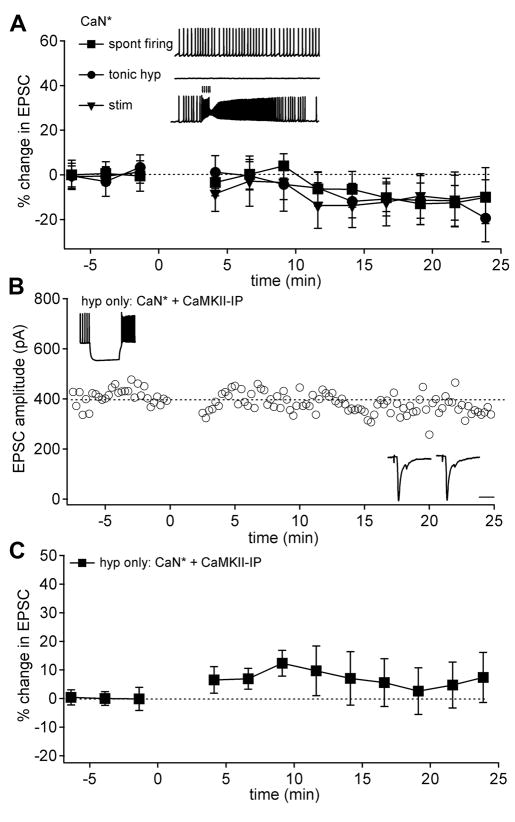
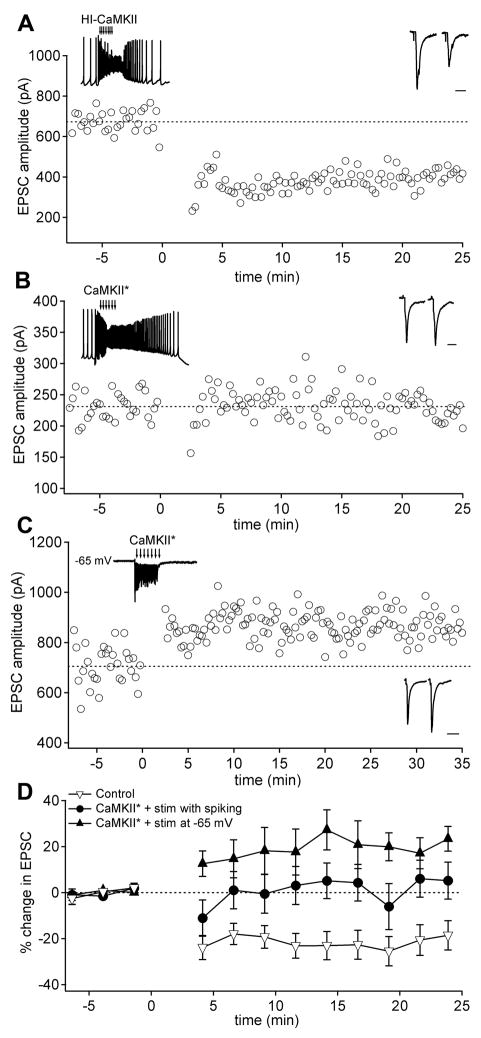
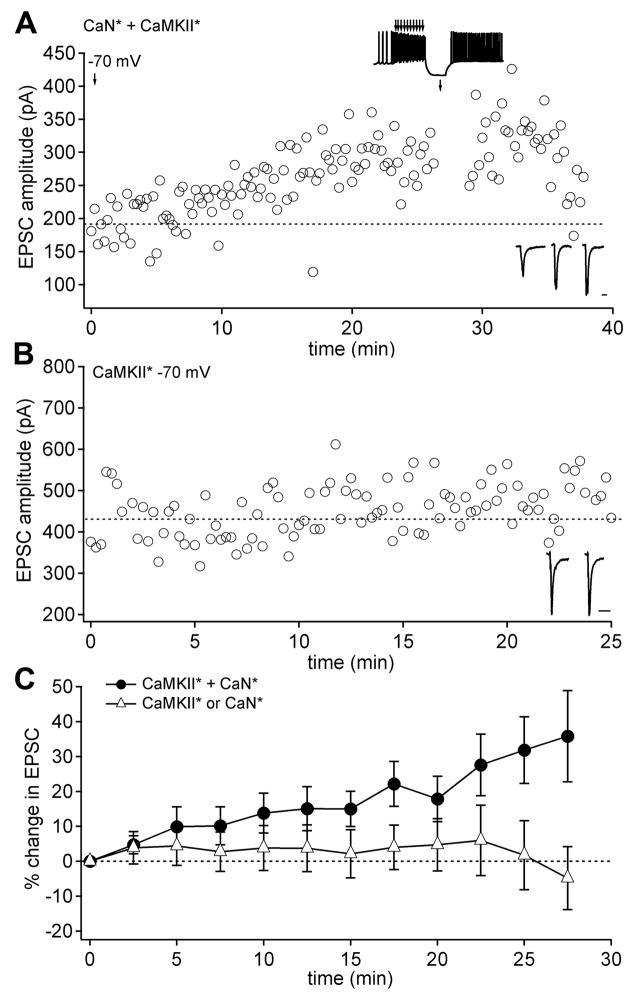
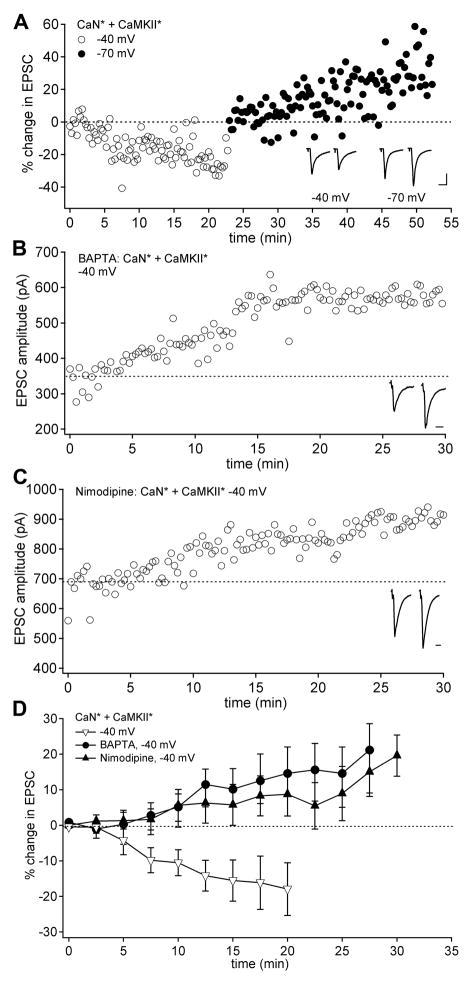
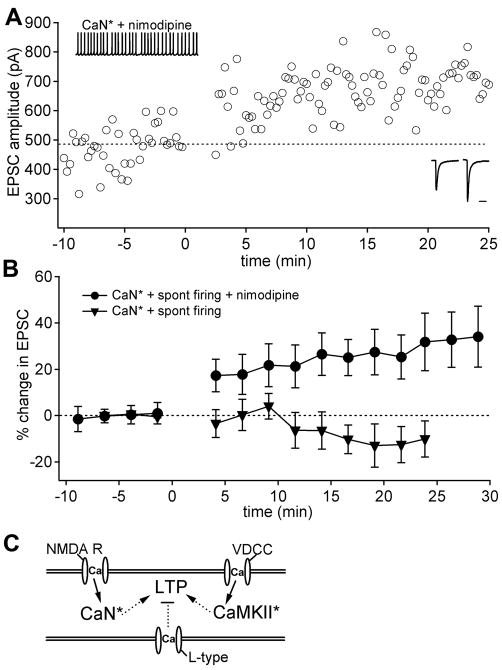
Long-term potentiation (LTP) of mossy fiber EPSCs in the cerebellar nuclei is controlled by synaptic inhibition from Purkinje neurons. EPSCs are potentiated by a sequence of excitation, inhibition, and disinhibition, raising the question of how these stimuli interact to induce plasticity. Here, we find that synaptic excitation, inhibition, and disinhibition couple to different calcium-dependent signaling pathways. In LTP induction protocols, constitutively active calcineurin can replace synaptic excitation, and constitutively active alpha-CaMKII can replace calcium influx associated with resumption of spiking upon disinhibition. Additionally, nimodipine can replace hyperpolarization, indicating that inhibition of firing decreases Ca influx through L-type Ca channels, providing a necessary signal for LTP. Together, these data suggest that potentiation develops after a calcineurin priming signal combines with an alpha-CaMKII triggering signal if and only if L-type Ca current is reduced. Thus, hyperpolarization induced by synaptic inhibition actively controls excitatory synaptic plasticity in the cerebellar nuclei.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Mechanisms of potentiation of mossy fiber EPSCs in the cerebellar nuclei by coincident synaptic excitation and inhibition.J Neurosci. 2008 Oct 15;28(42):10549-60. doi: 10.1523/JNEUROSCI.2061-08.2008. J Neurosci. 2008. PMID: 18923031 Free PMC article.
-
Ca currents activated by spontaneous firing and synaptic disinhibition in neurons of the cerebellar nuclei.J Neurosci. 2009 Aug 5;29(31):9826-38. doi: 10.1523/JNEUROSCI.2069-09.2009. J Neurosci. 2009. PMID: 19657035 Free PMC article.
-
Potentiation of mossy fiber EPSCs in the cerebellar nuclei by NMDA receptor activation followed by postinhibitory rebound current.Neuron. 2006 Jul 6;51(1):113-23. doi: 10.1016/j.neuron.2006.05.021. Neuron. 2006. PMID: 16815336
-
Synaptic inhibition, excitation, and plasticity in neurons of the cerebellar nuclei.Cerebellum. 2010 Mar;9(1):56-66. doi: 10.1007/s12311-009-0140-6. Cerebellum. 2010. PMID: 19847585 Free PMC article. Review.
-
Nothing can be coincidence: synaptic inhibition and plasticity in the cerebellar nuclei.Trends Neurosci. 2009 Mar;32(3):170-7. doi: 10.1016/j.tins.2008.12.001. Epub 2009 Jan 27. Trends Neurosci. 2009. PMID: 19178955 Free PMC article. Review.
Cited by
-
Modeling memory consolidation during posttraining periods in cerebellovestibular learning.Proc Natl Acad Sci U S A. 2015 Mar 17;112(11):3541-6. doi: 10.1073/pnas.1413798112. Epub 2015 Mar 3. Proc Natl Acad Sci U S A. 2015. PMID: 25737547 Free PMC article.
-
Bidirectional plasticity gated by hyperpolarization controls the gain of postsynaptic firing responses at central vestibular nerve synapses.Neuron. 2010 Nov 18;68(4):763-75. doi: 10.1016/j.neuron.2010.09.025. Neuron. 2010. PMID: 21092864 Free PMC article.
-
Multiple types of cerebellar target neurons and their circuitry in the vestibulo-ocular reflex.J Neurosci. 2011 Jul 27;31(30):10776-86. doi: 10.1523/JNEUROSCI.0768-11.2011. J Neurosci. 2011. PMID: 21795530 Free PMC article.
-
Calcium-based dendritic excitability and its regulation in the deep cerebellar nuclei.J Neurophysiol. 2013 May;109(9):2282-92. doi: 10.1152/jn.00925.2012. Epub 2013 Feb 20. J Neurophysiol. 2013. PMID: 23427305 Free PMC article.
-
The cerebellar nuclei take center stage.Cerebellum. 2011 Dec;10(4):633-6. doi: 10.1007/s12311-010-0245-y. Cerebellum. 2011. PMID: 21279491 Free PMC article. No abstract available.
References
-
- Aizenman CD, Linden DJ. Regulation of the rebound depolarization and spontaneous firing patterns of deep nuclear neurons in slices of rat cerebellum. J Neurophysiol. 1999;82:1687–1709. - PubMed
-
- Aizenman CD, Manis PB, Linden DJ. Polarity of long-term synaptic gain change is related to postsynaptic spike firing at a cerebellar inhibitory synapse. Neuron. 1998;21:827–835. - PubMed
-
- Aizenman CD, Linden DJ. Rapid, synaptically driven increases in the intrinsic excitability of cerebellar deep nuclear neurons. Nat Neurosci. 2000;3:109–111. - PubMed
-
- Anchisi D, Scelfo B, Tempia F. Postsynaptic currents in deep cerebellar nuclei. J Neurophysiol. 2001;85:323–331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
