

doi: 10.1016/j.cell.2010.05.019.
Quantitative genetic interactions reveal biological modularity
Affiliations
- PMID: 20510918
- PMCID: PMC2946632
- DOI: 10.1016/j.cell.2010.05.019
Item in Clipboard
Quantitative genetic interactions reveal biological modularity
Cell.
.
Abstract
Traditionally, research has been reductionist, characterizing the individual components of biological systems. But new technologies have increased the size and scope of biological data, and systems approaches have broadened the view of how these components are interconnected. Here, we discuss how quantitative mapping of genetic interactions enhances our view of biological systems, allowing their deeper interrogation across different biological scales.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
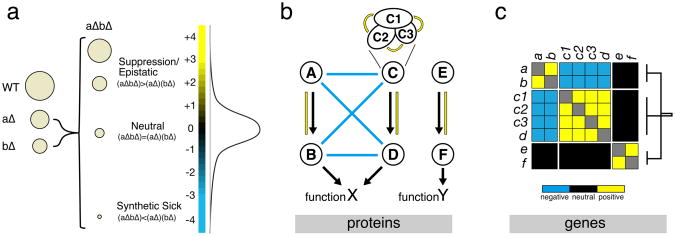
Definition and interpretation of genetic interactions (A) The spectrum of genetic interaction scores, as determined by typical E-MAP screens, ranges from negative (e.g. synthetic sickness) when fitness of the double mutant is less than expected, to positive (e.g. suppression) when the double mutant fitness is higher than expected. As exemplified by the distribution of genetic interaction scores, most gene pairs have genetic interaction scores close to zero (i.e. neutral). (B) Representation of an idealized example pathway. In this pathway, the components E and F are required for function Y. Components A through D are important for function X, although the AB branch is redundant with the CD branch. Additionally, C is an idealized 3 subunit complex. Both X and Y functions are important for viability but they are independent, and consequently no epistatic interactions exist between them. (C) Matrix of genetic interactions for example pathway. In this scenario, the branches AB, CD and EF are expected to be enriched for positive interactions within each cluster. Additionally, redundant branches as the AB and CD would typically show an enrichment of negative interactions between them. Neutral genetic interactions are expected between the EF branch and the ABCD module given their independent contributions to fitness. The genetic interaction scores of each gene with all others forms a phenotypic vector that can be analyzed using clustering methods. Hierarchical clustering of the expected genetic interaction scores for this example pathway is expected to result in three clearly distinct clusters (AB, CD and EF). We note that genetic interactions alone would not distinguish between the C complex subunits (c1, c2 and c3) from the additional branch component D.
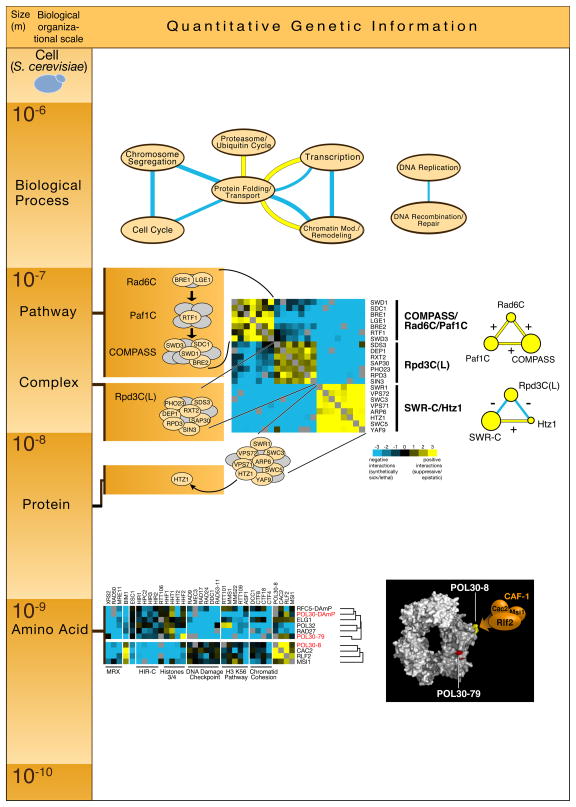
Quantitative genetic information obtained at different scales of biological organization. We use, as an example, data obtained from one E-MAP focused on chromatin biology (Collins et al., Nature, 2007) and exemplify how it can be used to reveal novel biological insight at different orders of magnitude of spatial resolution. At the top of the figure, genetic association between different processes was identified by searching for significant enrichment of positive (yellow edges) or negative (blue edges) genetic interactions between the processes listed. In the middle of the figure, hierarchical clustering of the genetic interaction obtained in this E-MAP revealed a module composed of three highly correlated clusters that are enriched in positive genetic interactions within each cluster and negative genetic interactions across them: 1) Rad6C, Paf1C and COMPASS, 2) Rpd3C(L) and 3) SWR-C/Htz1. At the bottom of the figure, hierarchical clustering of the two different point mutants (pol30-8 and pol30-79) of PCNA (Pol30) reveals different functions for different regions of the protein. See text for details of these biological findings derived from the quantitative genetic interaction data.
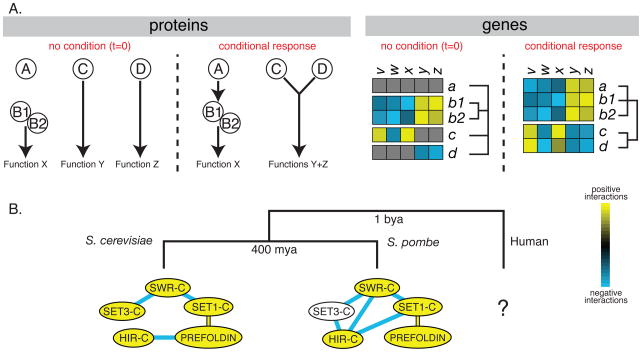
Condition and species dependent genetic interactions. (A) Changes in cellular interaction networks and corresponding genetic profiles after a change in external conditions. In our hypothetical example, a specific stimulus results in the activation of A which regulates complex B1/B2 that is important for Function X. This conditional dependent regulation is expected to be reflected in a conditional increase in the number of genetic interactions observed for gene a that should result in a profile that is highly correlated with those of b1 and b2. Changes in external conditions of our hypothetical example also result in post-translational modification in protein C that can now interact with protein D creating a conditional multifunctional complex. In this scenario, the profiles that are distinct in standard conditions become highly correlated after treatment with the stimulus. (B) Changes in genetic interaction networks over millions of years after speciation events. As species diverge through neutral and adaptive evolution, they accumulate changes that are expected to result in differences in their genetic interaction networks. We illustrate this divergence with the observed changes in genetic interactions between known protein complexes of S. cerevisiae and S. pombe, two species that have diverged from each other more than 400 million years ago (mya). The complexes with significant enrichment of positive genetic interactions within their complex subunits are denoted in yellow with blue and yellow edges connecting complexes that have a significant enrichment of negative or positive interactions, respectively, between the different modules. The illustration is based on experimental data obtained for both of these species using the E-MAP approach (Collins et al., 2007; Roguev et al., 2008). As with comparative genomics, understanding the evolution of genetic interactions will become clearer once more species are analyzed in the same way. We anticipate that tools currently under development will soon allow for the study of human cells that are expected to show even higher divergence given the larger divergence time to fungal species (~1 billion years – bya).
References
-
- Alber F, Dokudovskaya S, Veenhoff LM, Zhang W, Kipper J, Devos D, Suprapto A, Karni-Schmidt O, Williams R, Chait BT, et al. Determining the architectures of macromolecular assemblies. Nature. 2007;450:683–694. - PubMed
-
- Aloy P, Russell RB. Structural systems biology: modelling protein interactions. Nat Rev Mol Cell Biol. 2006;7:188–197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
